English
| 妛廗婰壇僾儘僕僃僋僩 | 岞塿嵿抍朄恖丂搶嫗搒堛妛憤崌尋媶強 妛廗婰壇偲偦偺忈奞偺暘巕婡峔偺夝柧僾儘僕僃僋僩 仹156-8506 搶嫗搒悽揷扟嬫忋杒戲2-1-6 揹榖丗03-6834-2425 |
|---|
尋媶撪梕偺徯夘乮徻嵶儁乕僕乯乮尋媶撪梕偺徯夘儁乕僕偵栠傞乯僔儑僂僕儑僂僶僄扨棧擼偵偍偗傞僉僲僐懱僔僫僾僗揱払偺 壜慪揑曄壔 暥愑丂忋栰峩暯/陰摗丂幚 偼偠傔偵 擋偄偲揹婥僔儑僢僋傪慻傒崌傢偣偨僔儑僂僕儑僂僶僄偺擋偄寵埆妛廗偼丄揟宆揑側屆揟揑忦審晅偗偱偁傞 (Quinn et al., 1974)丅堚揱妛傪慻傒崌傢偣偨徻嵶側尋媶偐傜丄偙偺忦審晅偗偵偼僉僲僐懱偲屇偽傟傞悢愮屄偺僯儏乕儘儞偐傜側傞堦懳偺恄宱廤抍偑廳梫側摥偒傪偟偰偄傞偙偲偑柧傜偐偵偝傟偰偒偨丅乮恾忋抜乯丅偝傜偵丄嬤擭偺僀儊乕僕儞僌夝愅偵傛傝丄擋偄寵埆妛廗傪偟偨僴僄偱偼偦偺擋偄偵懳偡傞僉僲僐懱偺恄宱妶摦偑憹嫮偟偰偄傞偙偲偑傢偐偭偨 (Wang et al., 2008)丅偡側傢偪丄偙偺擋偄偵懳偡傞僉僲僐懱墳摎偺壜慪揑曄壔偑丄擋偄妛廗偺婎斦偲側偭偰偄傞偲峫偊傜傟傞丅変乆偼僉僲僐懱恄宱妶摦偺壜慪揑曄壔偺暘巕嵶朎妛揑側幚懱傪柧傜偐偵偡傋偔丄in vitro偵偍偄偰僉僲僐懱偺壜慪揑曄壔傪僔僫僾僗儗儀儖偱夝愅偱偒傞宯傪奐敪偡傞偙偲偵偟偨丅 侾丏庢傝弌偟偨擼偱偺婰壇宍惉丠 in vitro偱偺夝愅偑恄宱妶摦偺壜慪揑曄壔偺棟夝偵桳岠偱偁傞偙偲偼丄傎擕椶偺奀攏僗儔僀僗傗傾儊僼儔僔偺攟梴偟偨姶妎丒塣摦僯儏乕儘儞偲偄偭偨in vitro幚尡宯偑妛廗婰壇尋媶偵懡戝側峷專傪偟偰偒偨偙偲偐傜傕柧傜偐偱偁傞丅嬶懱揑偵偼丄in vivo偲堘偄恄宱傊偺巋寖傪幚尡幰偑帺桼偵僐儞僩儘乕儖偱偒傞偙偲傗丄栻棟妛揑側夝愅偑梕堈側偙偲丄偝傜偵偼僀儊乕僕儞僌夝愅偺戝偒側忈奞偲側傞昗杮偺乽摦偒乿偑慡偔柍偄偙偲偵偁傞丅 僴僄偺拞悤恄宱宯傪扨棧偡傞偲丄擋偄偺堦師拞悤偱偁傞怗妎梩乮antennal lobe, AL乯偲揹婥僔儑僢僋傪擼偵揱偊傞懱惈姶妎拞悤偺暊悜恄宱愡乮ventral nerve code, VNC乯偑業弌偡傞丅偦偙偱AL偲丄擼-VNC娫傪宷偖慇堐乮ascending fibers of VNC, AFV乯傪奺乆僈儔僗揹嬌偱巋寖偡傞偙偲偱丄扨棧偟偨擼偵偁偨偐傕擋偄偲揹婥僔儑僢僋偑梌偊傜傟偰偄傞偐偺傛偆側巋寖傪梌偊傜傟傞偺偱偼側偄偐偲峫偊偨丅 僉僲僐懱偵Ca2+姶庴惈寀岝偨傫傁偔G-CaMP傪敪尰偝偣偨僴僄擼偱AL傪巋寖偡傞偲丄惗偒偨僴僄偱尒傜傟傞傛偆偵丄僉僲僐懱偺儘乕僽偲屇偽傟傞晹埵偱嫮偄Ca2+墳摎偑娤嶡偝傟偨乮恾拞抜嵍乯丅AL偲僉僲僐懱偼傾僙僠儖僐儕儞偵傛偭偰僔僫僾僗揱払偝傟偰偄傞偙偲偑抦傜傟偰偍傝丄幚嵺偵変乆偺宯偵偍偄偰傕僯僐僠儞惈傾僙僠儖僐儕儞庴梕懱乮nAChRs乯偺僽儘僢僇乕傪搳梌偡傞偲Ca2+墳摎偼徚幐偟偨丅 師偵丄AL偲AFV傪摨帪偵巋寖偡傞偲嫮偄Ca2+墳摎偑娤嶡偝傟乮恾拞抜拞乯丄偦偺屻嵞傃AL偩偗傪巋寖偡傞偲丄摨帪巋寖偡傞慜偵斾傋偰Ca2+墳摎偑憹嫮偟偰偄傞偙偲偑娤嶡偝傟偨乮恾拞抜塃乯丅傑偨偙偺憹嫮偱偼乮侾乯AL偲AFV偺摨帪巋寖偑偦偺桿摫偵廳梫偱偁傝丄AL偩偗傪嫮偔巋寖偡傞丄傕偟偔偼AL偲AFV傪暿乆偵巋寖偡傞曽朄偱偼桿摫偝傟側偄乮楢崌惈乯丄乮俀乯AL巋寖偵懳偡傞Ca2+墳摎惈偼憹嫮偡傞偑丄AFV巋寖偵懳偡傞Ca2+墳摎惈偼憹嫮偟側偄乮擖椡摿堎惈乯丄乮俁乯憹嫮偼丄2帪娫埲忋曐帩偝傟傞偑丄孞傝曉偟AL傪巋寖偡傞偲徚幐偡傞乮曐帩摿惈乯偲偄偭偨丄擋偄寵埆妛廗偵傛傞婰壇偲嬌傔偰傛偔椶帡偟偨摿挜偑傒傜傟偨丅懄偪擋偄寵埆妛廗偱傕乮侾乯擋偄偲揹婥僔儑僢僋偼摨帪偵梌偊傜傟傞昁梫偑偁傝丄乮俀乯忦審晅偗屻傕揹婥僔儑僢僋偵懳偡傞婖旔惈偑曄壔偡傞偙偲偼側偄丅傑偨乮俁乯擋偄寵埆妛廗婰壇偼悢帪娫埲忋曐帩偝傟傞偑丄忦審晅偗偵梡偄偨擋偄傪孞傝曉偟僴僄偵梌偊傞偲婰壇偺徚嫀偑婲偒傞丅偙偺憹嫮偼AL-MB娫偺僔僫僾僗揱払偺槾恑偵埶傞傕偺偱偁傝丄埲忋偺摿挜偐傜僴僄偺擋偄寵埆妛廗偺嵶朎妛揑儌僨儖偱偁傞偲峫偊丄偙傟傪奀攏偺LTP丄傾儊僼儔僔偺LTF偵曧偭偰丄long-term enhancement乮LTE乯偲屇傫偱偄傞丅 俀丏LTE偺僔僫僾僗婡峔 擋偄寵埆妛廗偵偍偄偰廳梫側摥偒傪偡傞偲峫偊傜傟偰偄傞恄宱揱払暔幙庴梕懱偑侾宆僪乕僷儈儞庴梕懱乮D1R丄僴僄堚揱巕柤DopR乯偲NMDA宆僌儖僞儈儞巁庴梕懱乮NR丄僴僄堚揱巕柤dNR乯偱偁傞丅変乆偼偙傟傜偺庴梕懱偑LTE宍惉偵偍偄偰偳偺傛偆側婡擻傪偡傞偺偐傪夝愅偟偨丅偙傟傑偱偺峴摦幚尡夝愅偐傜丄D1R偑揹婥僔儑僢僋傪僉僲僐懱偵揱偊傞偺偱偼側偄偐偲峫偊傜傟偰偄傞丅偟偐偟D1R傪寚懝偟偨僴僄偵偍偄偰AFV巋寖偵懳偡傞僉僲僐懱偺墳摎惈傪應掕偟偨偲偙傠丄AFV巋寖偵懳偡傞僉僲僐懱偺Ca2+墳摎惈偼D1R曄堎懱偱偼惓忢偱偁偭偨丅堦曽丄NR偺慾奞嵻傪搳梌偡傞偲AFV巋寖偵懳偡傞僉僲僐懱恄宱墳摎惈偼尠挊偵慾奞偝傟偨丅偱偼丄LTE宍惉偦偺傕偺偵NR偲D1R偼昁梫側偺偩傠偆偐丅AL偲AFV偺摨帪巋寖帪偵偦傟偧傟偺庴梕懱偵懳偡傞慾奞嵻傪搳梌偡傞偲丄偄偢傟偺応崌傕LTE宍惉偼慾奞偝傟偨丅D1R偵偍偄偰偼曄堎懱傪梡偄偰傕摨條偺寢壥偑摼傜傟偨丅AL偲AFV傪摨帪巋寖偡傞偙偲偑LTE宍惉偵昁恵偱偁傞偙偲偲暪偣偰丄nAChR丄NR偍傛傃D1R偑妶惈壔偝傟傞偙偲偑LTE宍惉偵昁梫偱偁傞偙偲偑柧傜偐偲側偭偨 (Ueno et al., 2012)丅 崱屻偺揥奐 偙傟傑偱丄僴僄偺妛廗婰壇尋媶偼峴摦妛傕偟偔偼暘巕堚揱妛揑夝愅偵傛傞抦尒偑庡偱偁傝丄偦偺娫偵偁傞嵶朎惈暔妛揑側抦尒偑晄廫暘偱偁偭偨丅杮尋媶偵傛傞扨棧擼偺僉僲僐懱恄宱壜慪惈偼丄擋偄寵埆妛廗偲傕椙偔椶帡偟偰偍傝丄峴摦偲堚揱巕傪宷偖夝愅宯偵側傞偲婜懸偝傟傞丅 丂扨棧擼LTE偺夝愅偐傜丄崱夞偼揹婥僔儑僢僋忣曬偑D1R偩偗偱側偔丄NR偵傛偭偰傕揱払偝傟傞偺偱偼側偄偐偲偄偆偙偲傪帵嵈偟偨丅堦曽丄D1R偼恄宱妶摦傪夘偝偢偵僉僲僐懱傊偲忣曬傪揱払偟乮偍偦傜偔偼G僞儞僷僋傪夘偟偨cAMP忣曬揱払宱楬乯丄僉僲僐懱偺壜慪揑曄壔傪堷偒婲偙偟偰偄傞偲峫偊傜傟傞丅偟偐偟丄幚嵺偵AFV偐傜偺巋寖偑D1R妶惈壔偵昁梫廫暘側偺偐偲偄偆偙偲偼晄柧偱偁傞丅傑偨恾偵帵偟偨傛偆偵丄nAChRs丄NR偍傛傃D1R偺妶惈壔偑LTE宍惉偵昁恵偱偁傞偲偡傟偽丄偳偺傛偆側暘巕偑偙偺3偮偺妶惈壔傪専抦偱偒傞偺偩傠偆偐丅偦傟偲傕丄傕偭偲帪娫暘夝擻偺崅偄夝愅傪峴偊偽丄3偮偺庴梕懱偺妶惈壔偵偼弴彉偑偁傞偙偲偑尒弌偝傟傞偺偩傠偆偐丅尰嵼変乆偼僀儊乕僕儞僌夝愅媄弍傪嬱巊偟偰丄偙傟傜偺栤戣偵挧愴拞偱偁傞丅 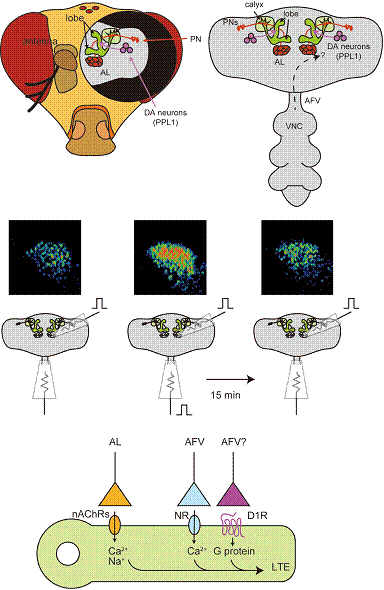 恾丂 扨棧擼僉僲僐懱偵偍偗傞恄宱壜慪惈 忋抜丗僴僄拞悤恄宱宯偺柾幃恾丅擋偄庴梕婍偱偁傞antenna偐傜庴梕嵶朎偑antennal lobe乮AL乯傊偲忣曬傪憲傝丄AL偐傜projection neurons乮PNs乯偵傛偭偰僉僲僐懱乮椢晹暘乯偺calyx傊偲忣曬偑揱払偝傟傞丅嫽暠偟偨僉僲僐懱偼偝傜偵lobe偲屇偽傟傞neuropile傊偲忣曬偑揱払偝傟傞丅揹婥僔儑僢僋側偳偵傛傝嫽暠偟偨ventral nerve code乮VNC乯偼ascending fiber偵傛偭偰擼傊偲忣曬傪揱偊傞丅尰嵼偱偼丄擋偄寵埆妛廗帪偺揹婥僔儑僢僋忣曬偼嵟廔揑偵PPL1偲屇偽傟傞僪僷儈儞嶌摦惈僯儏乕儘儞僋儔僗僞偵傛偭偰僉僲僐懱傊偲揱偊傜傟傞偲峫偊傜傟偰偄傞丅 拞抜丗寀岝幨恀偼偦傟偧傟偺巋寖捈屻偵偍偗傞lobe偱偺Ca2+墳摎傪媈帡僇儔乕偱帵偟丄柾幃恾偼巋寖偺曽朄傪帵偟偰偄傞丅扨棧偟偨擼偺AL傪僈儔僗揹嬌偱巋寖偡傞偲僉僲僐懱偺lobe偱Ca2+墳摎偑娤嶡偝傟傞乮嵍乯丅AL偲摨帪偵AFV傕巋寖偡傞偲嫮偄Ca2+墳摎偑娤嶡偝傟乮拞乯丄偦偺15暘屻偵嵞傃AL偩偗傪巋寖偡傞偲丄埲慜傛傝傕嫮偄墳摎惈偑娤嶡偝傟傞乮塃乯丅 壓抜丗崱夞偺尋媶偵傛傝悇掕偝傟傞僉僲僐懱僯儏乕儘儞乮椢晹暘乯偱偺壜慪揑曄壔偺暘巕宱楬丅AL巋寖偵傛傝nAChRs偑妶惈壔偟丄梲僀僆儞偑棳擖偡傞丅AFV巋寖偱偼NR偑妶惈壔偟丄Ca2+偑棳擖偡傞丅AFV巋寖偼摨帪偵D1R傕妶惈壔偡傞偲峫偊傜傟丄D1R偼G僞儞僷僋傪夘偟偰嵶朎撪忣曬揱払宱楬傪妶惈壔偡傞丅偙傟傜偺3偮偺斀墳偵傛傝丄LTE偑宍惉偝傟傞丅 Quinn, W.G., Harris, W.A., and Benzer, S. (1974). Conditioned behavior in Drosophila melanogaster. Proc Natl Acad Sci U S A 71, 708-712. Ueno, K., Naganos, S., Hirano, Y., Horiuchi, J., and Saitoe, M. (2012). Long-term enhancement of synaptic transmission between antennal lobe and mushroom body in cultured Drosophila brain. J Physiol. Wang, Y., Mamiya, A., Chiang, A.S., and Zhong, Y. (2008). Imaging of an early memory trace in the Drosophila mushroom body. J Neurosci 28, 4368-4376. 乮尋媶撪梕偺徯夘儁乕僕偵栠傞乯 |