最新ニュース
2026年
- 2026年 5月11日 東京都医学総合研究所にて第16回都医学研シンポジウム「ヒト脳進化を駆動した哺乳類脳の発生原理」を主催し、開催しました。(写真)
- 2026年3月、研修生の和田が新潟大学で開催された「新潟大学博士学生支援プログラムシンポジウム – 次世代を担う博士人材 – 博士イノベーターという新たな可能性」でポスター発表を行い、優秀研究ポスター賞を受賞しました。(写真)
- 2026年2月、研修生の土井(お茶の水女子大学)が卒業論文発表を終えて学士号を取得し、山崎(新潟大学大学院)および石塚(早稲田大学大学院)も修士論文発表を無事終えて修士号を取得しました。
2025年
- 2025年12月11〜12日 先進ゲノム支援2025年度拡大班会議で守屋研究員が口頭発表をし、丸山リーダーと隈元研究員がポスター発表をしました。
- 2025年12月3~5日 第48回日本分子生物学会年会にて、高沢研究員、研修生の宋、山崎、篠原がポスター発表をしました。
- 2025年11⽉10⽇〜11⽇ AMED国際シンポジウム2025「The Early-Life Nexus」(慶應義塾⼤学 三⽥キャンパス)にて、丸⼭リーダーが招待講演を、研修⽣杉⽥、⼭崎がポスター発表を⾏いました。
- 2025年10⽉22⽇ 所内で開催した2025年度研修⽣等発表会で研修⽣の杉⽥、⼭崎が発表を⾏い、杉⽥が最優秀賞を受賞しました。(写真、写真2 )
- 2025年9月11~13日 名古屋で開催された第68回日本神経化学会大会で研修生の和田・杉田が発表を行い、杉田が優秀発表賞を受賞しました。(写真)
- 2025年7月30~8月1日 高校生を対象とした夏のセミナーを開催し実習をしました。(写真)
- 2025年7月24~27日 第48回日本神経科学大会に丸山リーダーが参加し、畠中研究員、研修生の山崎・篠原がポスター発表をしました。(写真)
- 2025年7月14~16日 英国オックスフォード大学St. John’s Collegeにて行われたThe Anatomical Society Summer Meetingに参加し、丸山リーダーが口頭発表、研修生の阿知波がポスター発表をしました。(写真)
- 2025年6月27~28日 AMED-CREST「健康・医療の往生に向けた早期ライフステージにおける生命現象の解明」領域 丸山班「サブプレート神経活動のダイナミズムによる脳神経回路の発達機構」第4回研究班会議を東京都医学総合研究所の講堂にて行いました。(写真)
- 2025年6月4~8日 Black Sea Neurogenesis 2025(ブルガリア)にて、丸山リーダーが口頭発表をしました。(写真)
- 2025年6月2~3日 英国・オックスフォード大学のSherrington Libraryにて、丸山リーダーが神経科学テーマのゲストスピーカーとして講演を行いました。(写真)
- 2025年5月25~28日 Cortical Development Conference 2025(シチリア)にて丸山リーダーが口頭発表をし、守屋・松村研究員と研修生の杉田がポスター発表を行い、杉田がBest Question賞を受賞しました。(写真)
- 2025年 4月15~16日 東京都医学総合研究所にて第30回国際シンポジウム「Principles of Neocortical Development and Evolution II」を主催し、開催しました。(写真)
- 2025年4月 原・元ゲノム研究センター研究員と隈元研究員、丸山リーダーらは、「The spatial transcriptome of the late-stage embryonic and postnatal mouse brain reveals spatiotemporal molecular markers」についてScientific Reportsに発表しました。
- 2025年4月 野村客員研究員と丸山リーダーは「Genetic and developmental bases for mammalian neocortical evolution」についてDevelopment, Growth & Differentiationに発表しました。
- 2025年 4月 3~5日 Brain Development Symposium Kyoto 2025(京都女子大学主催)にて、Zoltan Molnar教授と丸山リーダーが講演を行いました。(写真)
- 2025年3月 Zoltan Molnar教授が都医学研セミナーで講演を行いました。(写真)
- 3月10日:「Development and Evolution of Thalamocortical Connectivity」
- 3月27日:「Transient Circuits in the Developing Brain」
- 2025年3-4月 Zoltan Molnar教授が解剖学講義を3回シリーズで行いました。(写真)
- 3月13日:「Lectures on neuroanatomy-I: Thomas Willis (1621-1675) The Founder of Neuroanatomy and Clinical Neurology」
- 3月26日:「Lectures on neuroanatomy-II: Insight into the Life and Work of Sir Chales Sherrington(1857-1952)」
- 4月9日:「Lectures on neuroanatomy-III: Evolutionary Developmental Biology of the Mammalian Cerebral Cortex」
- 2025年3月9日〜5月7日まで、英国・オックスフォード大学のZoltan Molnar教授を招聘しました。(写真)
- 2025年1~3月にかけて研修生(野村・お茶の水女子大学大学院、片山・杉田・早稲田大学大学院、宋・都立大学大学院)の修士論文発表がそれぞれの大学院で無事終わり、修士号を取得しました。
- 2025年3月12日 所内研究発表会第3部会で優秀発表者賞を研修生の野村が受賞しました。(写真)
- 2025年2月22日 東京都医学総合研究所の2024年度都医学研第8回都民講座「脳とはどのようにして生まれ、進化してきたのか?―ヒト脳への進化の道のりと病―」にて、丸山リーダーが「脳とは何か、どのようにできるのか?」の講演を行いました。(写真)
- 2025年 1月16~17日 AMED領域班会議(TKPガーデンシティ博多)にて、丸山リーダーが口頭発表(オンライン)を、松村研究員がポスター発表を行いました。
- 2025年 1月28日 第14回日本マーモセット研究会で守屋研究員がポスター発表をしました。
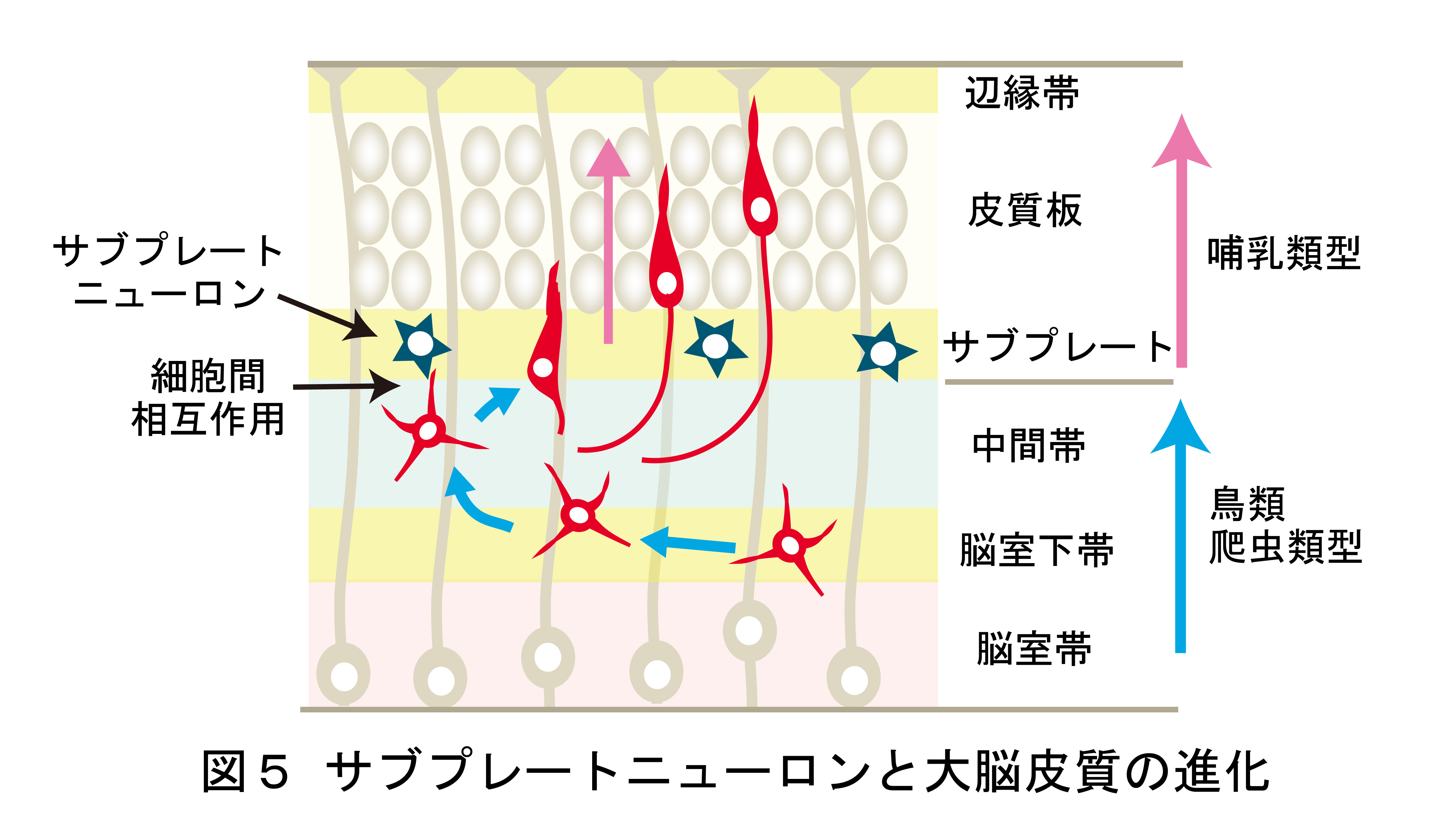
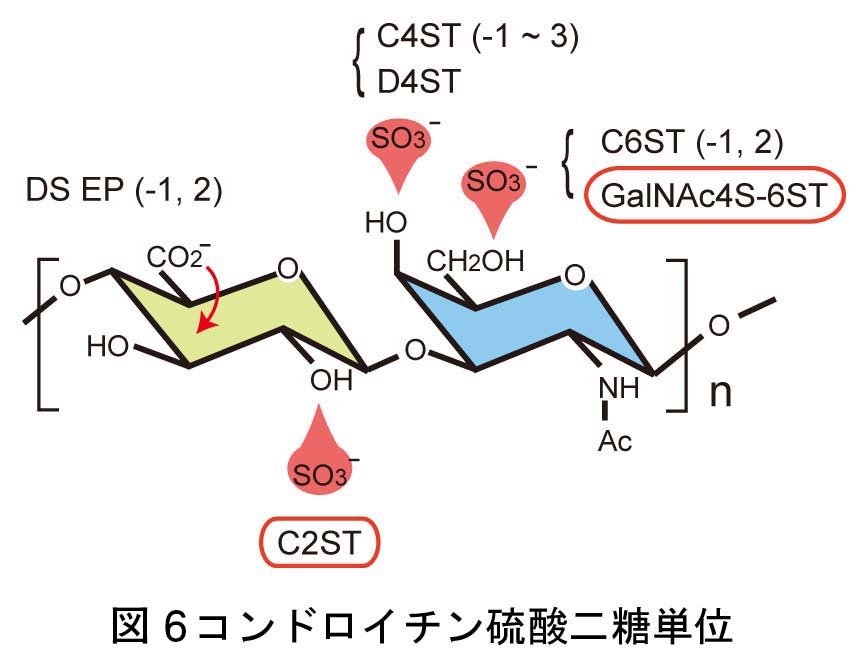
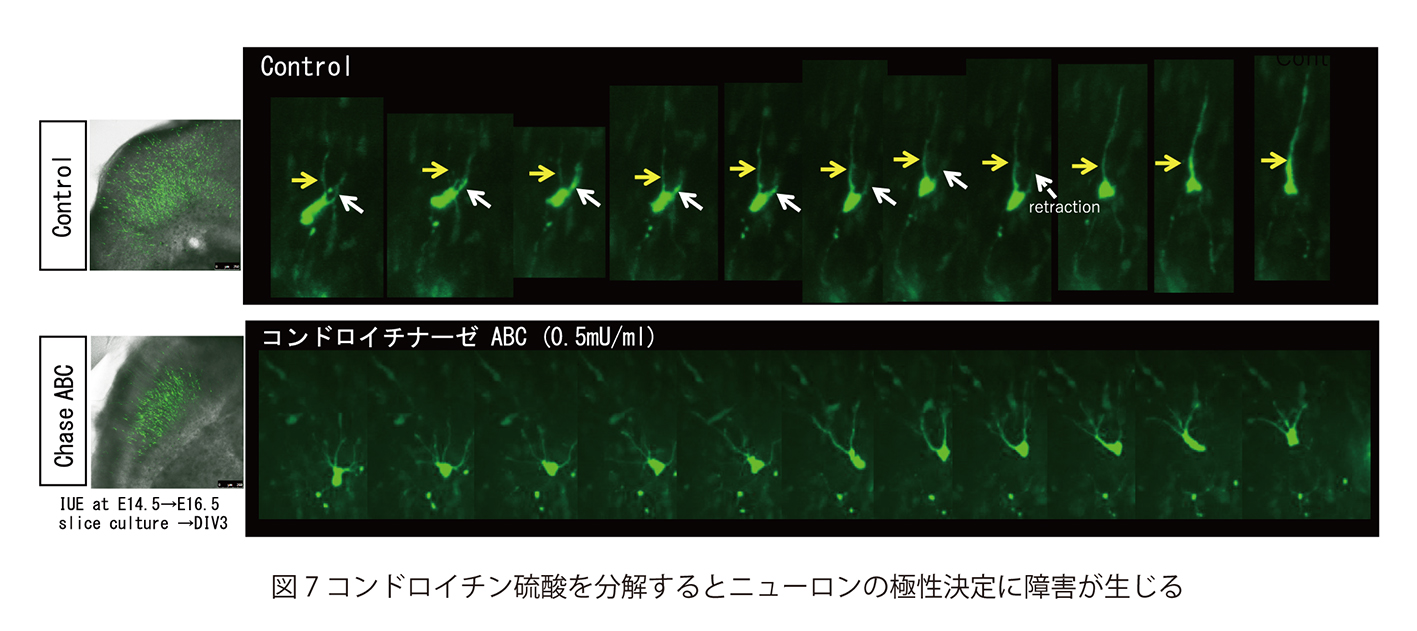
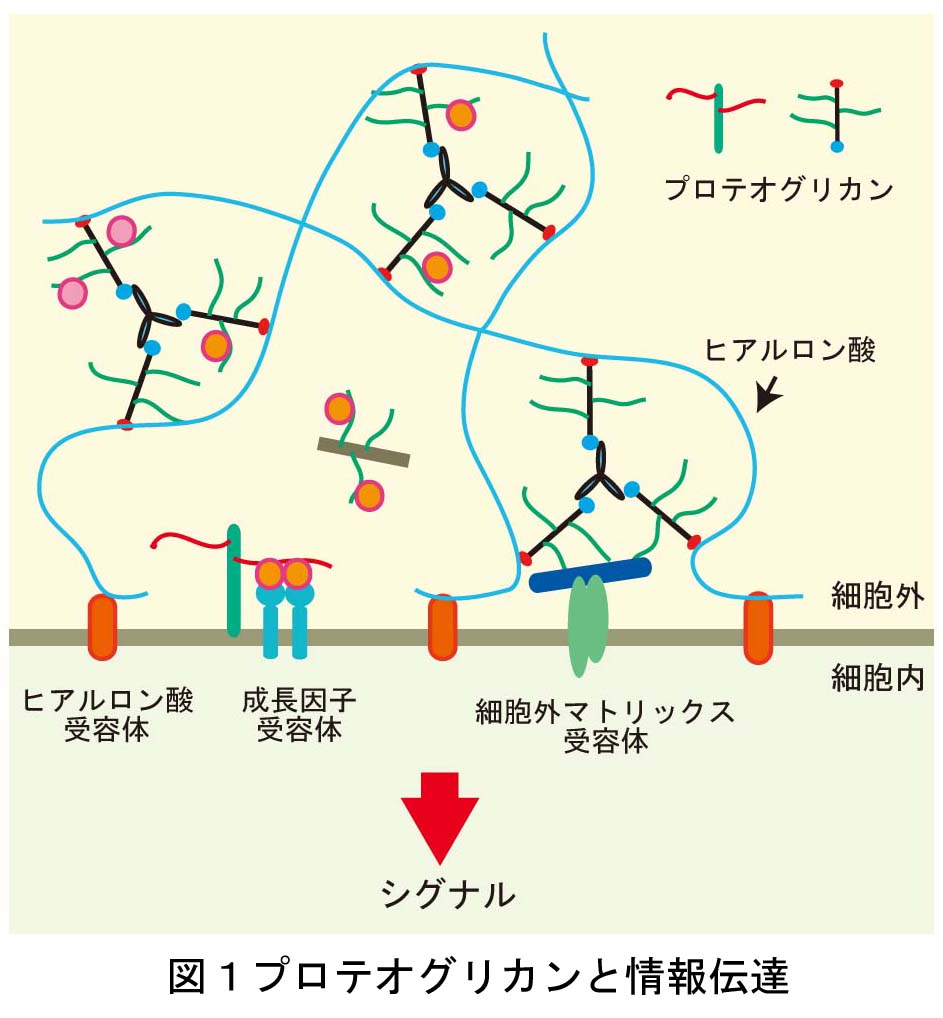
脳神経系を構成する精緻な神経回路は、
どのようなメカニズムで形成されるのか?
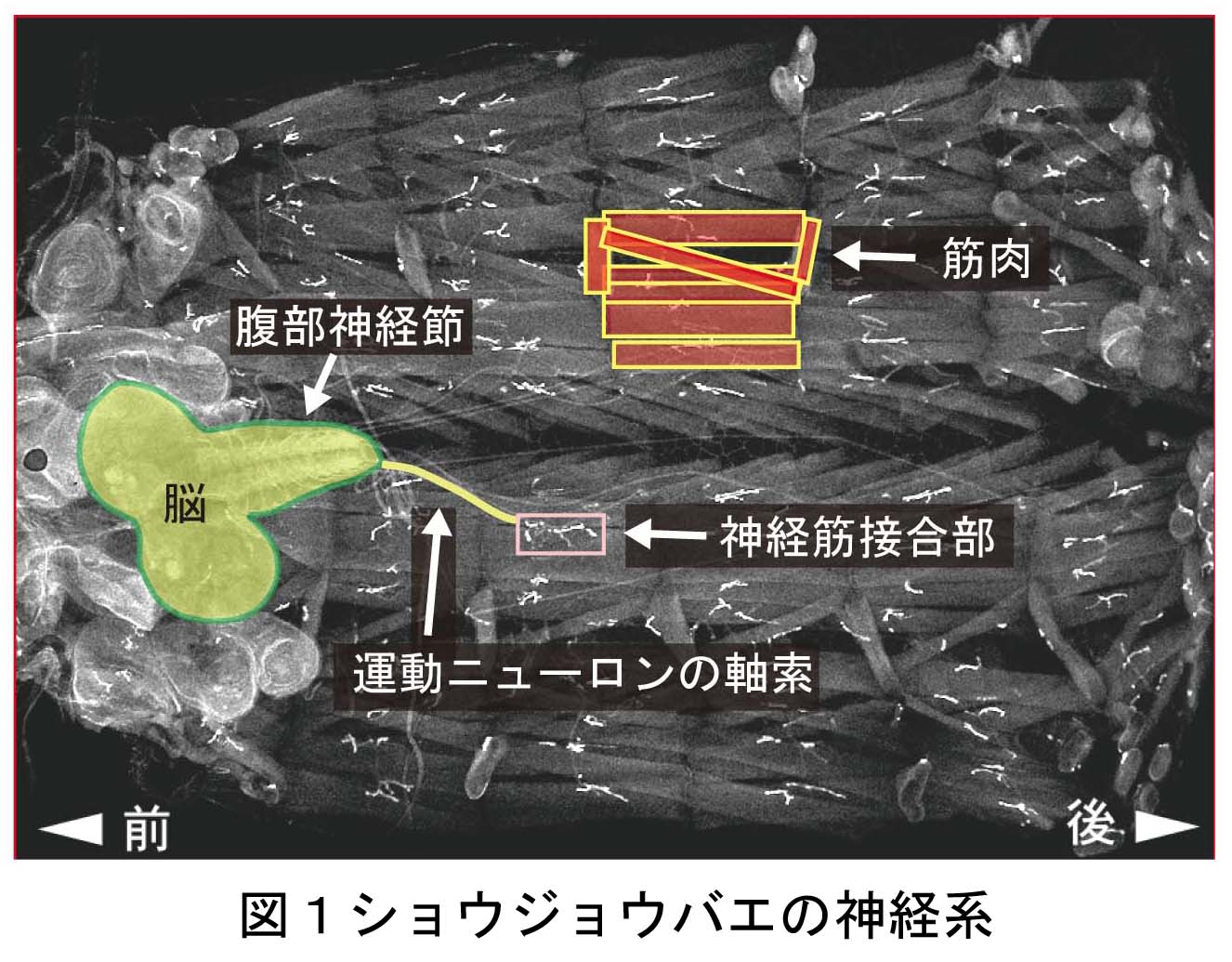
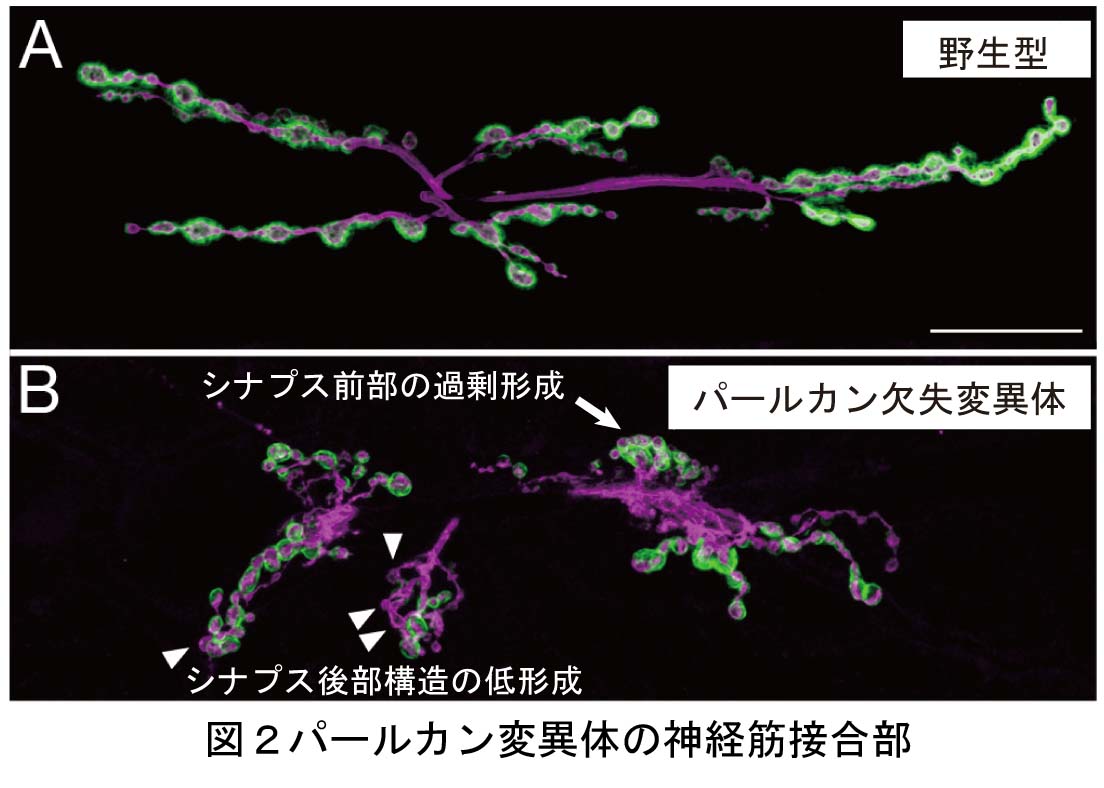
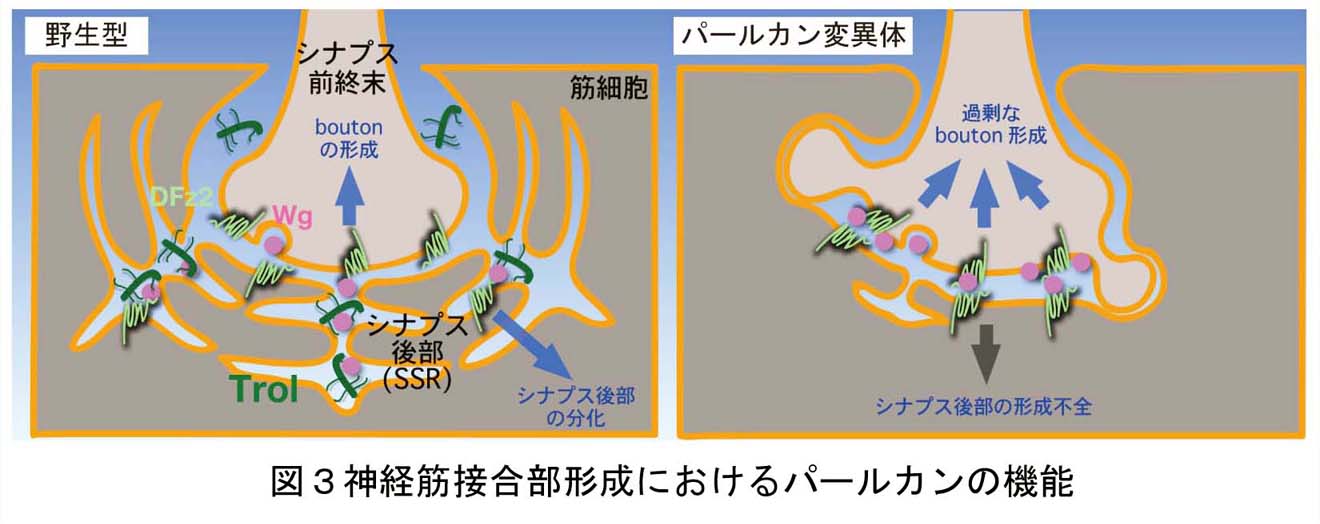
これは、今なお新たな課題が次々に立ち現われてくる深い問題です。神経回路形成は、複雑かつ多様な細胞-細胞間、及び細胞-細胞外マトリックス間の相互作用の下に進行します。本プロジェクトでは、マウス大脳皮質及びショウジョウバエ神経筋接合部をモデルに、このような相互作用の分子メカニズムを解明することを目指しています。