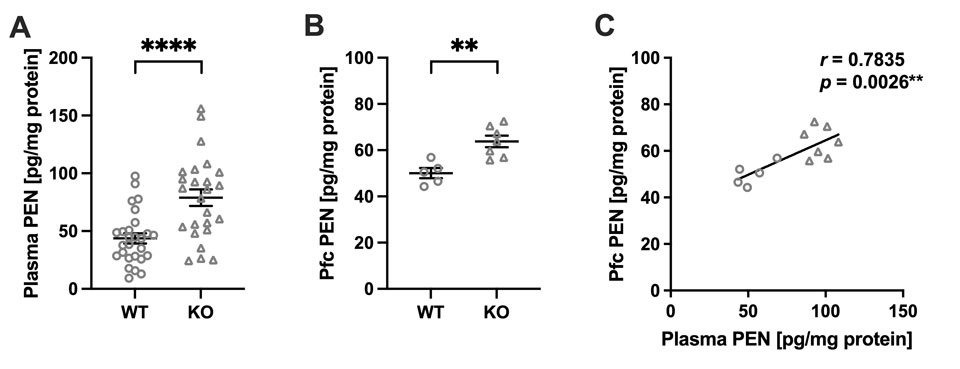
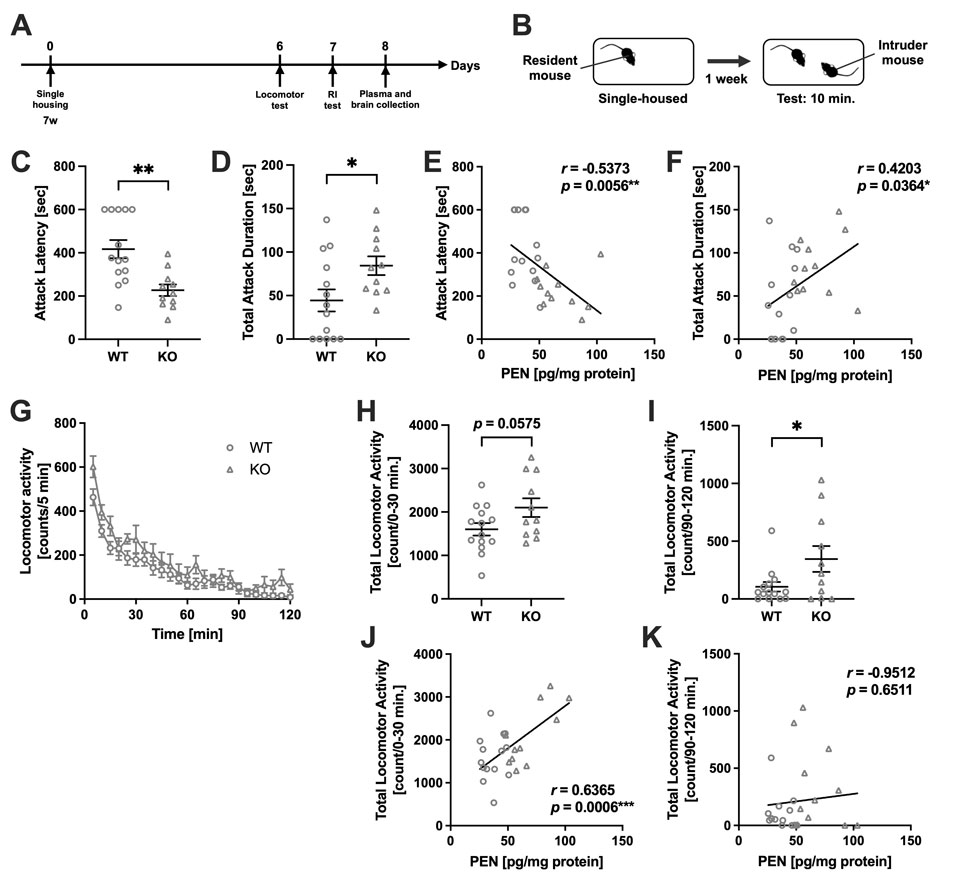
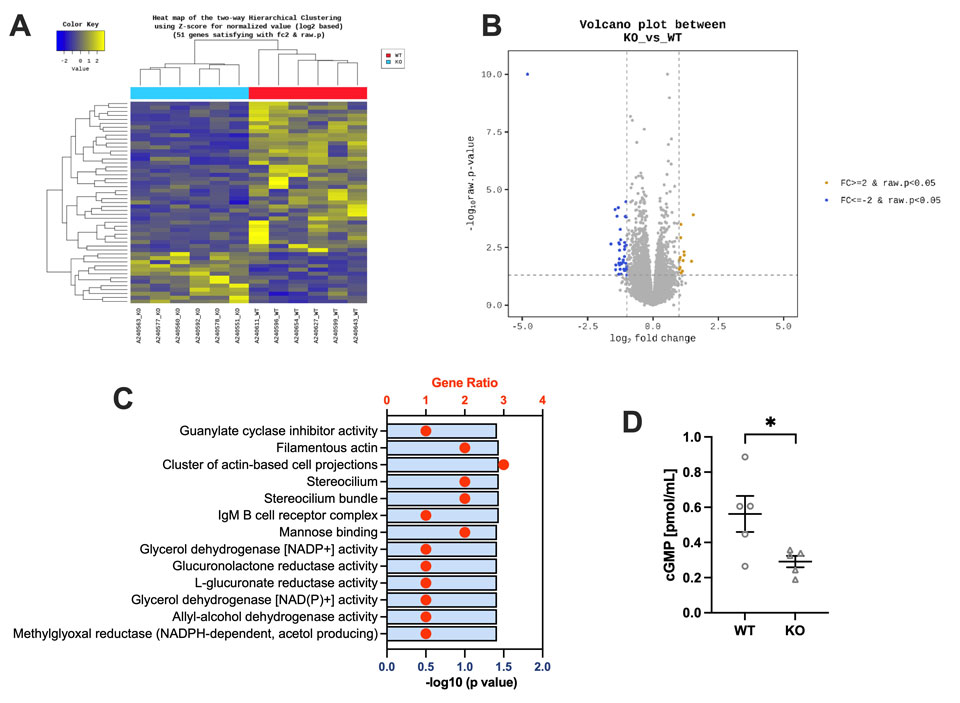
Akr1a欠失により誘導されるペントシジン蓄積が攻撃性亢進および過活動を引き起こす
Pentosidine accumulation induced by Akr1a deficiency drives aggression and hyperactivity
25
K. Iino, K. Toriumi, M. Miyashita, K. Suzuki, K. Tabata, S. Miyata, M. Takahashi, J. Fujii, M. Itokawa, M. Arai, Pentosidine accumulation induced by Akr1a deficiency drives aggression and hyperactivity, Neurosci. Lett. 877 (2026) 138557.
doi.org/10.1016/j.neulet.2026.138557
J. Chaudhuri, Y. Bains, S. Guha, A. Kahn, D. Hall, N. Bose, A. Gugliucci, P. Kapahi, The role of advanced glycation end products in aging and metabolic diseases: bridging association and causality, Cell Metab. 28 (2018) 337-352. https://doi.org/10.1016/j.cmet.2018.08.014
N. Rabbani, M. Xue, P.J. Thornalley, Dicarbonyls and glyoxalase in disease mechanisms and clinical therapeutics, Glycoconj. J. 33 (2016) 513-525. https://doi.org/10.1007/s10719-016-9705-z
N. Rabbani, P.J. Thornalley, Protein glycation – biomarkers of metabolic dysfunction and early-stage decline in health in the era of precision medicine, Redox Biol. 42 (2021) 101920. https://doi.org/10.1016/j.redox.2021.101920
A. Goldin, J.A. Beckman, A.M. Schmidt, M.A. Creager, Advanced glycation end products: sparking the development of diabetic vascular injury, Circulation 114 (2006) 597-605. https://doi.org/10.1161/CIRCULATIONAHA.106.621854
M. Arai, H. Yuzawa, I. Nohara, T. Ohnishi, N. Obata, Y. Iwayama, S. Haga, T. Toyota, H. Ujike, M. Arai, T. Ichikawa, A. Nishida, Y. Tanaka, A. Furukawa, Y. Aikawa, O. Kuroda, K. Niizato, R. Izawa, K. Nakamura, N. Mori, D. Matsuzawa, K. Hashimoto, M. Iyo, I. Sora, M. Matsushita, Y. Okazaki, T. Yoshikawa, T. Miyata, M. Itokawa, Enhanced carbonyl stress in a subpopulation of schizophrenia, Arch. Gen. Psychiatry 67 (2010) 589-597. https://doi.org/10.1001/archgenpsychiatry.2010.62
M. Miyashita, M. Arai, H. Yuzawa, K. Niizato, K. Oshima, I. Kushima, R. Hashimoto, M. Fukumoto, S. Koike, T. Toyota, H. Ujike, T. Arinami, K. Kasai, M. Takeda, N. Ozaki, Y. Okazaki, T. Yoshikawa, N. Amano, T. Miyata, M. Itokawa, Replication of enhanced carbonyl stress in a subpopulation of schizophrenia, Psychiatry Clin. Neurosci. 68 (2014) 83-84. https://doi.org/10.1111/pcn.12081
M. Miyashita, M. Arai, A. Kobori, T. Ichikawa, K. Toriumi, K. Niizato, K. Oshima, Y. Okazaki, T. Yoshikawa, N. Amano, T. Miyata, M. Itokawa, Clinical features of schizophrenia with enhanced carbonyl stress, Schizophr. Bull. 40 (2014) 1040-1046. https://doi.org/10.1093/schbul/sbt129
K. Toriumi, K. Iino, A. Ozawa, M. Miyashita, S. Yamasaki, K. Suzuki, H. Sugawa, K. Tabata, S. Yamaguchi, S. Usami, M. Itokawa, A. Nishida, R. Nagai, H. Kamiguchi, M. Arai, Glucuronic acid is a novel source of pentosidine, associated with schizophrenia, Redox Biol. 67 (2023) 102876. https://doi.org/10.1016/j.redox.2023.102876
K. Iino, K. Toriumi, R. Agarie, M. Miyashita, K. Suzuki, Y. Horiuchi, K. Niizato, K. Oshima, A. Imai, Y. Nagase, I. Kushima, S. Koike, T. Ikegame, S. Jinde, E. Nagata, S. Washizuka, T. Miyata, S. Takizawa, R. Hashimoto, K. Kasai, N. Ozaki, M. Itokawa, M. Arai, AKR1A1 variant associated with schizophrenia causes exon skipping, leading to loss of enzymatic activity, Front. Genet. 12 (2021) 762999. https://doi.org/10.3389/fgene.2021.762999