ドーパミン研究
ドーパミンによる精神疾患への関与および、行動の制御に関する研究
ドーパミンは神経伝達物質の一つで、人格、運動、動機づけなど、極めて重要な脳機能を担っています。加えて、統合失調症や注意欠陥多動性障害(ADHD)などの精神疾患とも深く関わっています。 私たちは、ドーパミンが疾患とどのように関連するのか、またどのような機序で行動制御を担っているのかを明らかにするために、ドーパミントランスポーターノックアウトマウス(DATKOマウス)と、ドーパミン欠乏マウス(DDマウス)を用いて研究を行っています。
DATKOマウスの研究
DATは、神経細胞の終末の前シナプスから放出されたドーパミンを、細胞内に再取り込みする働きをしています。 DATKOマウスではドーパミンが取り込まれず、細胞外のドーパミン濃度が高くなり、ドーパミン神経伝達が亢進します。 これに起因して、DATKOマウスは、多動、注意欠如、学習障害、衝動性など顕著なADHD様の行動を示します。
DATKOマウスにADHDの治療に広く用いられているメチルフェニデート(MPH)を投与すると、多動が抑制されましたが、これに対し、野生型マウスにMPHを投与すると、活動量が亢進しました。このことは、ADHD患者ではMPHによって多動が抑えられるのに対して、健常者では逆に活動量が亢進することと類似しています。 また、多動だけでなく、DATKOマウスで認められる学習障害も、MPHによって改善することが見出されました。(図1)
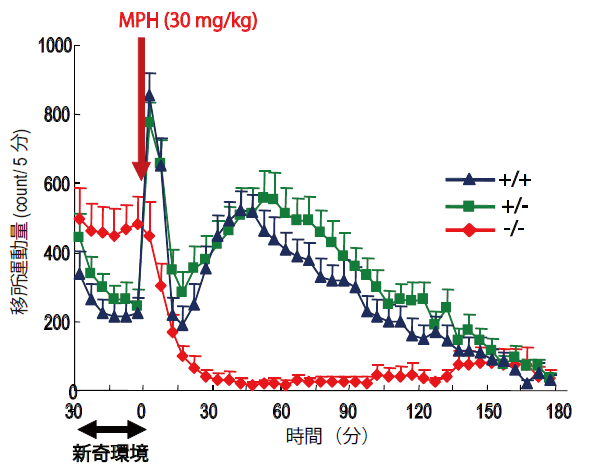
MPHはDATおよびノルエピネフリントランスポーター(NET)を阻害する作用を有しています。運動制御に関わる線条体では、DATがドーパミン再取り込みを担っています。 したがって、野生型マウスでは、MPHでDATが阻害されると、線条体ドーパミン量が上昇し、活動量の亢進が引き起こされます。 これに対し、DATKOマウスではDATがないため、MPHを投与しても線条体のドーパミン量は変化しません。一方、前頭前野では、DATが発現しておらず、NETが主にドーパミン再取り込みを担っているため、野生型、DATKOマウスともにMPHによってドーパミン量が上昇します。 前頭前野でドーパミン神経伝達が亢進することが、MPHによるDATKOマウスのADHD様行動の改善と関係しているのではないかと考えられます。
このように、DATKOマウスはADHDのとてもよいモデル動物で、これを用いることで病態メカニズムの解明や、治療薬開発を目指した研究を行っています。
DDマウスの研究
DDマウスは、ドーパミンやアドレナリンなどのカテコラミンの合成酵素の一つであるチロシン水酸化酵素の遺伝子を欠損させ、さらにアドレナリン神経細胞やノルアドレナリン神経細胞でチロシン水酸化酵素を発現するように遺伝子を組み換えたマウスです。 このマウスはドーパミンを作ることができないため、通常は生後2週齢ほどで死んでしまいますが、ドーパミン前駆体であるL-ドーパを毎日投与することで生き続けることができます。
成獣まで維持したDDマウスに対して、L-ドーパ投与を中止すると、24時間後では脳内にはまだドーパミンはわずかに残存していますが、72時間後には検出限界以下のレベルまで欠乏します。 L-ドーパ投与中止24時間後、72時間後に新奇環境下における活動量を調べたところ、24時間後では活動量の著しい低下がみられましたが、72時間後では予想外なことに、活動量の亢進が認められました(図2)。
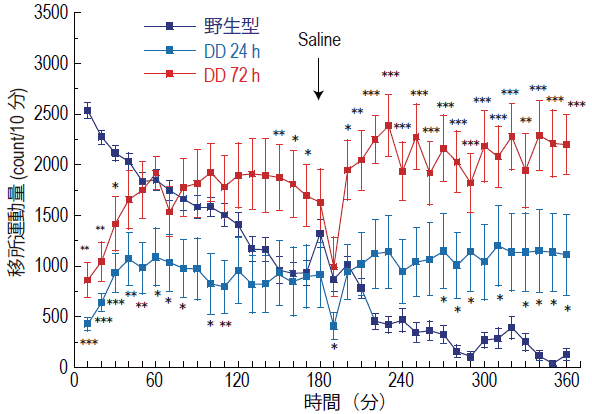
活動量の亢進は、非定型抗精神病薬の一つであるクロザピンで抑えられることがわかりました。クロザピンの作用は多岐にわたりますが、その中でもアセチルコリン系を活性化させる作用が、活動量の抑制に関わっていることがわかりました。
実際に、DDマウスの線条体ではアセチルコリン濃度の低下およびアセチルコリン合成酵素であるコリンアセチルトランスフェラーゼの発現の減少が観察されており、アセチルコリン系の変化が活動量の増加に関係する可能性が考えられます。
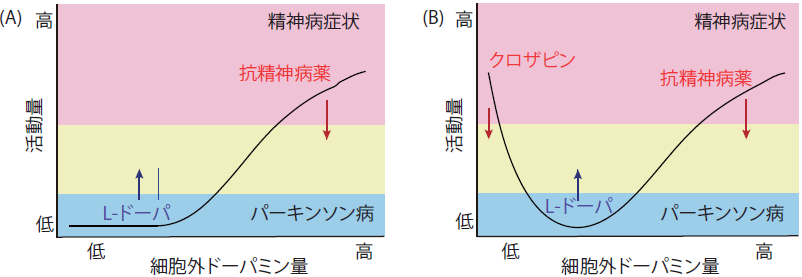
ドーパミン量と活動量とは、従来は比例関係にあると考えられてきましたが(図3A)、DDマウスの研究からドーパミン量が極端に減少すると活動量が亢進することが見出されました(図3B)。 ドーパミンが極端に減少した際の行動とそのメカニズムを解明し、ドーパミンがどのようにして行動の制御を行うのか、その真の機序を明らかにすることを目指した研究を行っています。