医学・生命科学全般に関する最新情報





![]()


![]()
ウイルスは、通常、自分自身がコードするタンパク質機能は限定されており、大部分、宿主の機能に依存して複製し、増殖します。したがって、SARS-CoV-2感染時に関与する宿主因子、あるいは細胞内経路の全貌を明らかにすることは、抗ウイルス剤の開発にとって重要です。最近、米国ロックフェラー大学を中心とした研究グループによりCell誌に発表された二報の論文で、SARS-CoV-2の増殖に必要とされる、タンパク質や細胞内経路が新たに発見されました。
最初の論文では、ゲノムワイドCRISPRノックアウトスクリーニングにより、SARS-CoV-2及び、3種類の季節性コロナウイルス (HCoV-OC43, HCoV-NL63, 及び HCoV-229E)の増殖に必要な遺伝子や経路を同定しました。
この実験ではCas9を発現するHuh-7.5肝細胞癌細胞(Huh-7.5- Cas9)を用いました。この細胞は内在性にSARS-CoV-2受容体(ACE2; angiotensin-converting enzyme 2)及び膜貫通型セリンプロテアーゼ(TMPRSS2; transmembrane serine protease 2)を発現しており、SARS-CoV-2、HCoV-OC43、HCoV-NL63及びHCoV-229Eウイルスに感染し、死滅します。研究グループはsgRNAsライブラリーを導入し、薬剤で選択し、その後、ウイルスを感染させ、生き残ってくる細胞を選択しました。ウイルス感染やウイルスによる細胞死に要求される遺伝子が破壊された時には、細胞が生存することが期待されます。生存細胞が有するsgRNAを解析することにより、ウイルス増殖に関与する宿主遺伝子を同定することができます。
その結果、グリコサミノグリカン(glycosaminoglycan)生合成、ステロール制御因子結合タンパク質(sterol regulatory element-binding protein; SREBP)シグナリング、骨形成タンパク質(bone morphogenetic protein; BMP)シグナリング、及びグリコシルフォスファチジルイノシトール(glycosylphosphatidylinositol)合成の経路が関与することが明らかになりました。さらに、いくつかの解析がされていない未知のタンパク質の要求性も明らかになりました。
解析の結果、VMP1、TMEM41B、及びTMEM64(VTT)ドメイン含有膜貫通型 タンパク質41(TMEM41B)が、SARS-CoV-2及び、3種類の季節性コロナウイルスの感染に必須であることが示されました。
もう一方の論文では、ヒトに脊髄炎、髄膜脳炎、脳炎を起こすフラビウイルス科ウイルスの複製にTMEM41B及びVMP1が要求されることを、こちらも、CRISPR-Cas9機能喪失スクリーニングにより発見しました。著者らは、フラビウイルスが感染し、ウイルスタンパクが合成されると、TMEM41Bが小胞体膜にウイルスタンパク質NS4A及びNS4Bとともにrecruitされ、複製複合体が集合し、ウイルス複製が進行すると提唱しています。興味深いことに、TMEM41B遺伝子にIle266Val/Leu置換のSNPが見出されました。このSNPは、野生型に比較して、フラビウイルス感染効率を低下させ、1/10以下の量のウイルスしか産生することができません。さらに、ヨーロッパやアフリカではほとんど検出されませんが、ラテンアメリカでは3.5%、東南アジア諸国では何と20%の頻度で検出されました。
上述のようにTMEM41BはSARS-CoV-2の感染にも必須です。TMEM41Bに存在するこのSNPは、SARS-CoV-2の感染、複製にも影響を与える可能性があります。したがって、TMEM41Bは東南アジア諸国で、新型コロナウイルスの感染数が低いことの一因となっている可能性があります。
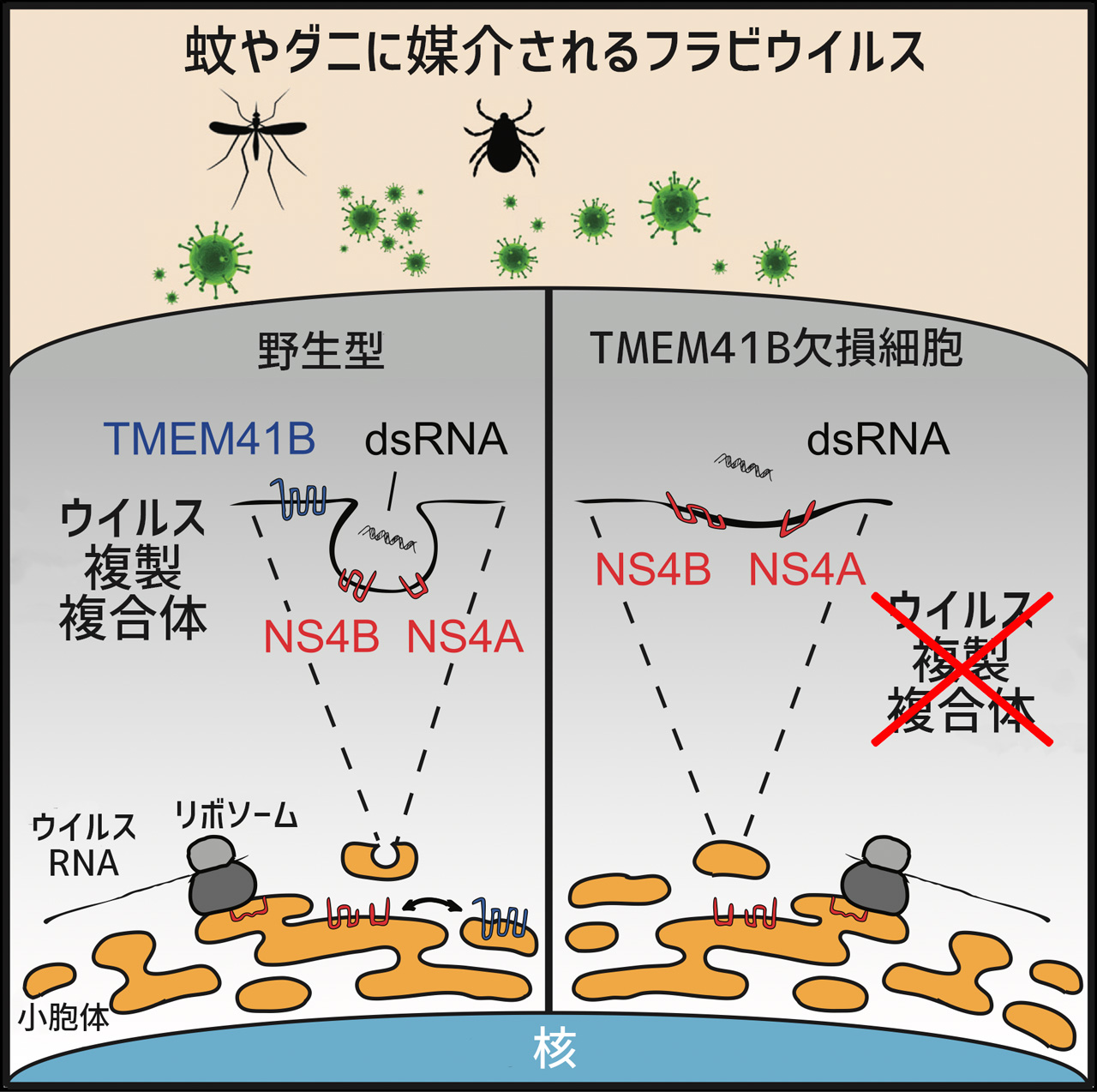
図. 小胞体の膜に存在するTMEM41Bタンパク質はフラビウイルス由来のNS4A, NS4Bタンパク質とともにウイルスRNA複製のために必要な複合体を形成する。TMEM41Bが存在しない場合には、複製複合体が適切に形成されないため、RNA複製が進行しない。TMEM41Bは同様なメカニズムでSARS-CoV-2 ゲノムRNAの複製にも関与している可能性がある。