蛋白質代謝研究室 吉田 雪子
私たちの身体を構成する細胞内のタンパク質には、それぞれ固有の寿命があります。プロテアソームは、タンパク質の寿命を制御するとともに、損傷を受けたタンパク質や合成に失敗した異常タンパク質を分解するタンパク質分解装置です。異常タンパク質が蓄積すると、アルツハイマー病やパーキンソン病など、加齢に伴って発症する神経変性疾患を引き起こすことが知られています。そのため、プロテアソーム活性が正常に保たれることは非常に重要です。ユビキチンという小さなタンパク質が、標的タンパク質のリジン残基に鎖状に連なる「ユビキチン鎖」を形成し、これがプロテアソームによる分解の目印となるという機序の発見に、ノーベル化学賞が授与されています。
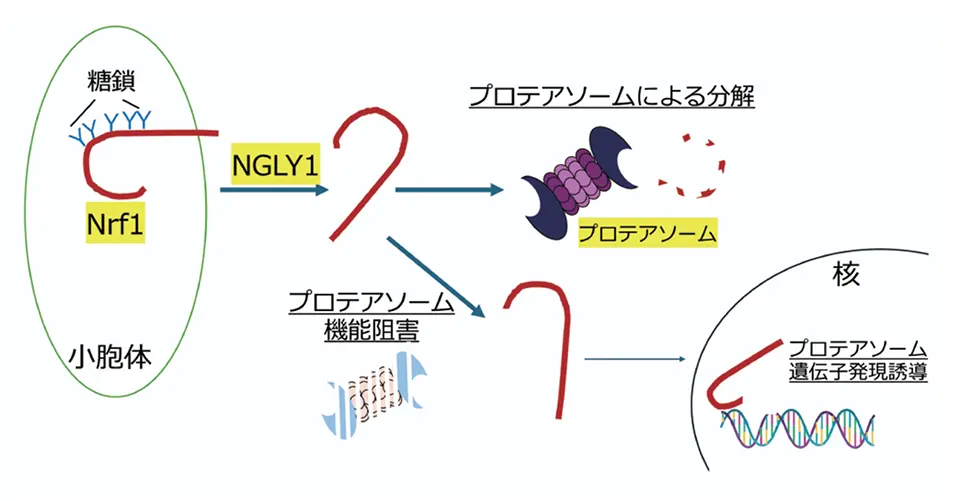
プロテアソーム活性が低下すると、Nrf1 という糖鎖を持つタンパク質の働きによって新たなプロテアソームが合成されるようになります。Nrf1 はプロテアソーム活性が正常な時はプロテアソームによって分解されるため細胞にはほとんど存在しません。プロテアソーム活性が低下した時にだけ、分解されずに核に移動してプロテアソームの合成を促す「転写因子」として働きます(図1)。転写因子として働くためには NGLY1 という糖鎖除去酵素が Nrf1の糖鎖を取り除く必要があることが、NGLY1 遺伝子に変異を持つ希少疾患「NGLY1 欠損症」の研究を通じてわかってきました。
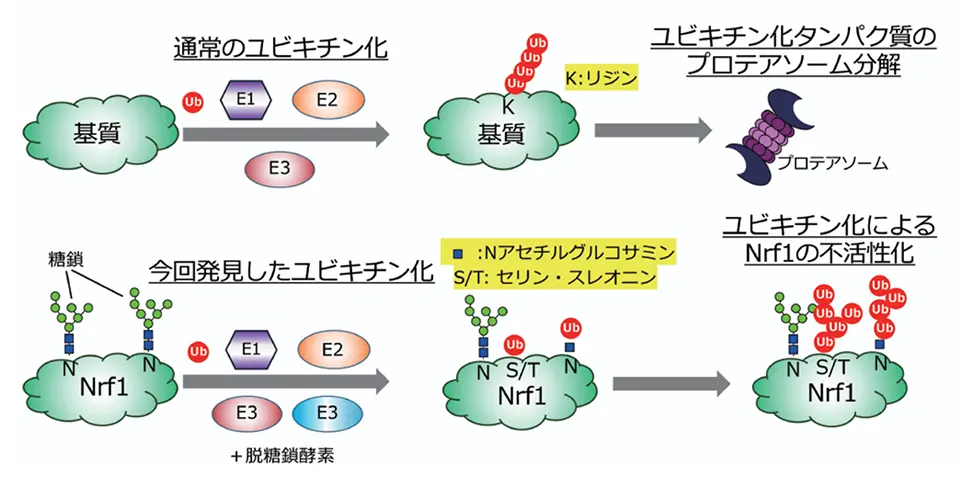
今回私たちは、NGLY1 がない細胞ではユビキチン鎖がついた Nrf1 が、プロテアソームに分解されずに細胞の中に溜まってしまうこと、さらに、この Nrf1 はプロテアソームを合成できないどころか、プロテアソームにダメージを与えてしまうことに気づきました。これが NGLY1 欠損症の原因であるのではないかと考え、Nrf1 についたユビキチン鎖の性質を明らかにすべく研究を進めました。その結果、このユビキチン鎖は通常のリジンにではなく、糖鎖の近くのセリンやスレオニンというアミノ酸や N アセチルグルコサミンという糖に結合している新たなタイプのユビキチン鎖であることがわかりました。ユビキチン化は通常1つの E3 ユビキチンリガーゼという酵素によって行われますが、この反応では Nrf1 の糖鎖を掴む E3 とそれに結合する別の E3 の2つの酵素がおこなっていることがわかりました。この特殊なユビキチン鎖がついた Nrf1 はプロテアソームによる分解を受けずに、しかし、ユビキチン鎖が核へ行くのを妨げるため、プロテアソーム活性を回復させられないことが疾患発症をひきおこすと考えられます(図2)。このユビキチン化を行う E3 を阻害する化合物は疾患の治療法の開発につながるものと期待されます。
Yoshida Y et al. Sugar-medicated non-canonical ubiquitination impairs Nrf1/NFE2L1 activation. Molecular Cell, 2024, 84 (16), 3115‒3127, doi: 10.1016/j.molcel.2024.07.013 (糖を介した非典型ユビキチン化は Nrf1/NFE2L1 の活性化を阻害する)
