

Oct. 2020 No.039
特集
Topics
開催報告
編集後記
Topics
知性の起源につながる細胞のかたちの変化
~大脳皮質の進化をもたらした神経細胞移動に関する論文が掲載~
脳神経回路形成プロジェクトリーダー丸山 千秋
研究の背景
哺乳類の大脳皮質では、神経細胞が移動モードを変化させながら脳の深部から表層へと移動し、6層構造ができますが、哺乳類独自の層構造が進化の過程でどのようにして出現したのかは謎に包まれていました。本研究では、哺乳類と爬虫類の大脳の発生過程における細胞移動様式を比較し、細胞のかたちの変化を制御するメカニズムがどのように進化してきたかの一端を解明しました。
哺乳類は爬虫類や鳥類と同じ羊膜類に属していますが、爬虫類の大脳はサイズが小さく、3層のシンプルな構造を形成することから、爬虫類の背側皮質から哺乳類の大脳皮質が進化したのではないかと考えられてきました。しかしながら、哺乳類以外の皮質構造の発生過程の研究は皆無でした。
本研究の概要
哺乳類大脳皮質の発生過程では、神経細胞の移動様式は3種類知られています。神経前駆細胞が突起を縮めながら細胞体が表層に移動するモード(細胞体トランスロケーション)、複数の突起を様々な方向に伸ばしつつ移動するモード(多極性移動)、さらに双極性に変化し、神経前駆細胞の放射状突起を上って脳の表層に向かうモード(ロコモーション)です(図1)。新しく産生された神経細胞は移動を終えた細胞を乗り越えて脳の表層に到着する、いわゆる「インサイド-アウト」様式で6層構造が形成されます。
本研究では、爬虫類の皮質構造が形成される際の神経細胞の移動様式や形態変化について観察を行いました。ヤモリの胚を用いて移動中の細胞を観察すると、ロコモーション型の移動モードは観察されません(図1)。また、爬虫類の皮質は外側から内側へと神経細胞が蓄積していく「アウトサイド-イン」様式で形成されることが明らかになりました。
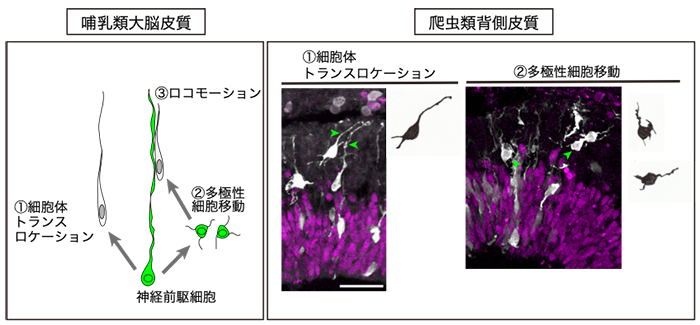
図1
哺乳類の大脳皮質と爬虫類背側皮質の発生過程における神経細胞移動。哺乳類大脳皮質(左)の形成過程では、神経前駆細胞から産生された神経細胞は①細胞体トランスロケーション、②多極性細胞移動、③ロコモーションの3つの移動モードによって大脳皮質を構築する。一方、爬虫類の背側皮質(右)では①、②の細胞移動は観察されたが、③ロコモーション型の細胞移動が観察されなかった。
次に、共通する移動モードと、種に固有の移動モードがどのような分子機構によって制御されているのかを検証しました。爬虫類(ヤモリ)の神経細胞を哺乳類(マウス)の大脳皮質に移植したところ、爬虫類の神経細胞はロコモーション型にはなりませんでした。
次にWnt(ウィント)シグナルに着目して調べ、マウスでは神経細胞移動様式の転換の際、細胞内のWntシグナルの活性化レベルが減弱することがわかりました。一方、爬虫類では、Wntシグナルが高レベルのまま維持されていました。そこで、爬虫類の神経細胞でWntシグナルを減弱させると、多極性から細胞体トランスロケーション型へとモードが変化した一方、ロコモーションモードは出現しなかったことから、このモードは哺乳類独自の発生メカニズムに依存していることが推測されました(図2)。
今後の展望
本研究は哺乳類の大脳皮質を構成する神経細胞の移動様式とその進化をもたらしたメカニズムの一端を明らかにし、ヒトの脳の成り立ちやその由来に対する新たな知見をもたらしました。研究成果は、脳の先天疾患の中でも神経細胞移動の異常に起因する疾患の原因に関する進化医学的な理解を与えるものです。なお本研究は、京都府立医科大学、理化学研究所との共同研究の成果です。
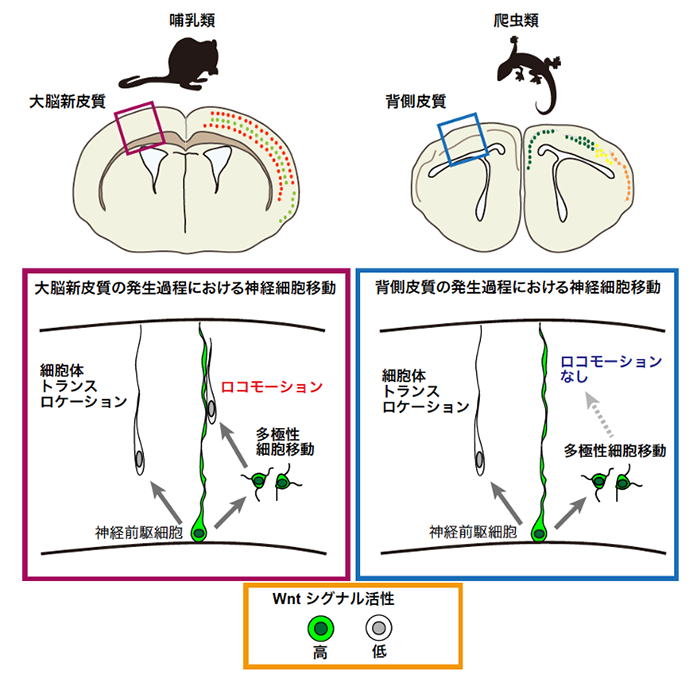
図2
哺乳類と爬虫類の脳の発生過程における神経細胞移動とWntシグナルの役割。多極性細胞移動ではWntシグナル活性が高い状態で維持されている。一方、細胞体トランスロケーション、ロコモーションの移動モードではWntシグナルは低い状態となる。ロコモーション型の神経細胞移動は哺乳類大脳皮質の発生過程でしか見られず、この細胞移動モードの獲得が大脳皮質の解剖学的特徴の進化をもたらしたと考えられる。