

脳は“傷つきながら”作られる
-ニューロン移動中にDNA損傷が発⽣するしくみを発⾒-
- 脳⽪質形成過程において、密な組織の中を移動するニューロンの核は圧迫され、中に含まれるDNA損傷が多発する
- DNA損傷はゲノムの中でも安全な領域に⽣じて速やかに修復され、ニューロンの⽣存や機能への影響は⼩さい
- 発⽣期にニューロンが受けたDNA損傷が残ると、神経疾患や⽼化の原因になりうる
京都⼤学アイセムス(⾼等研究院 物質―細胞統合システム拠点:WPI-iCeMS)の⾒學美根⼦教授、張喆菁研究員(当時:⽣命科学研究科⼤学院⽣)、⾼等研究院 ヒト⽣物学⾼等研究拠点:WPI-ASHBi の Andres Canela 特定研究員(当時:放射線⽣物研究センター特任准教授)らの研究グループは、当研究所ゲノム動態プロジェクトの笹沼博之プロジェクトリーダー、東京⼤学の岸雄介准教授、⼤阪⼤学の古⽥貴寛教授、シンガポール国⽴⼤学の Gianluca Grenci 博⼠らと共同で、正常な脳発⽣過程で、ニューロン遊⾛に伴う⼒学的ストレスにより DNA 損傷が多発することを発⾒しました。この成果は6⽉17⽇(英国時間)に Nature 誌でオンライン公開されました。
論文情報
- <論文タイトル>
- “Confined migration induces non-lethal DNA damage in developing neurons”
(参考訳:狭⼩環境での細胞遊⾛は発⽣期ニューロンに⾮致死的DNA損傷を誘導する) - <著者>
- 張喆菁, Andres Canela, 栗栖純⼦, Peilin Zou, 川上巧, 中澤直⾼, ⽵⽥理⼦, 佐伯⿇⾐, 宇都宮優希, Merve Bilgic, ⽯舘⽂善, Gianluca Grenci, 古⽥貴寛, 岸雄介, 笹沼博之, ⾒學美根⼦
- <発表雑誌>
- Nature
DOI: 10.1038/s41586-026-10648-8
URL:https://doi.org/10.1038/s41586-026-10648-8
<概要>
哺乳類の脳⽪質には、数百億のニューロンが整然と配置され、精密な神経回路を形成しています。脳⽪質構造は、幹細胞層で誕⽣したニューロンが密な神経組織を分け⼊って細胞移動(遊⾛)し、秩序正しく配列することで形成されます。狭い空間を遊⾛する際、ニューロンの核は激しく変形するため、核内の染⾊体にも⼒学的ストレスが加わると考えられます。
研究グループは、マウス⼩脳のニューロン遊⾛を最先端のライブイメージング技術で観察し、遊⾛中に圧迫された核内で DNA ⼆重鎖切断(DSB)が頻繁に⽣じていることを発⾒しました。⼀般的に DSB は細胞死や発ガンを誘発する最も重篤な DNA 損傷です。しかしニュ ーロンは DSB を多発しながらも細胞死を引き起こすことなく、個体の⽣涯を通じて正常に機能します。次世代 DNA シーケンサーを⽤いて全ゲノム上で DSB の発⽣部位を探索すると、遺伝⼦発現に影響する部位を避け、機能的影響の少ないゲノム上の不活性領域に形成されていることがわかりました。この過程には、DNA に捻りや張⼒のストレスがかかると応答してDSB を起こすトポイソゼラーゼが関与し、計画的に安全なゲノム領域が切断されるためであることが明らかになりました。また、ニューロン遊⾛中に⽣じた DSB は、半⽇以内に速やかに修復されることもわかりました。これらの結果は、ニューロンの遊⾛による⼒学的ストレスにより DSB が⽣じることは不可避である一方、それを安全に処理するメカニズムが実装されていることを⽰しています。
仮に脳発⽣中に⽣じた DSB が修復されずに残存した場合の影響を調べるため、DSB 修復酵素をニューロンのみで⽋損した変異マウスを作成しました。この変異マウスでは⼩脳ニュ ーロンに多くの DSB が蓄積するものの、細胞死は起こらず、外⾒上は正常に発育しました。しかし脳内の炎症細胞が活性化し、ニューロンの機能遺伝⼦の発現が低下していました。そして加齢に伴い、⼩脳機能の異常による運動障害が進⾏しました。これらの結果は、正常な発⽣過程においてニューロンで⽣じる DSB の修復エラーが、神経疾患の発症につながる潜在的リスクであることを⽰しています。
本研究により、ニューロンにおける⼒学的ストレスとゲノム安定性の新たな関係が明らかになりました。発⽣中の DSB 制御の破綻がゲノム不安定性疾患や⼀部の発達性障害の原因となる可能性が⽰唆され、これらの疾患の病因解明への重要な⼿掛かりとなることが期待されます。
1. 背景
細胞が組織内を⾃発的に移動する細胞遊⾛は、組織の発⽣、免疫応答、損傷治癒、ガン転移などの様々な⽣理的および病理的過程で⾒られます。染⾊体 DNA を内包する核は、遊⾛する細胞にとって最も⼤きく硬い積荷であり、狭い組織間隙を通過しようとすると核がつかえて前進できなくなります。そのため、遊⾛能(転移性)の⾼い浸潤性ガン細胞は、特殊な核膜分⼦構造※1 をもち、通常の体細胞より核が柔軟で、狭い空間でも⼤きく変形しながら通過できることが知られています。

⼀⽅で、遊⾛中の⼒学的ストレスを受けた核は、圧迫や変形によりしばしば傷つくことで、核膜の破損や DNA 損傷※2 を引き起こし、細胞死をもたらすことが報告されていました。こうした DNA 損傷が突然変異を誘発し、薬剤耐性や浸潤性の増⼤といったガンの悪性化に寄与する可能性も⽰唆されてきました。しかし、これらは実験室で株化した培養細胞※3 を⽤いた知⾒であり、⽣体内での細胞遊⾛で核の破損が起こっている様⼦を観察した例はほとんどなく、その⽣理的・病理的な意義については⼗分に解明されていませんでした。
哺乳類脳のニューロンは⾼い遊⾛能をもち、幹細胞からニューロンが誕生すると順序正しく遊⾛・配列することで脳⽪質を形成します。ニューロンの核も柔軟で、緻密な神経組織の狭い空間を⼤きく変形してすり抜けることができます。本研究により、驚くべきことに、正常な発⽣過程におけるニューロン遊⾛でも、狭い組織間隙を通過する際の⼒学的ストレスにより、DNA 損傷が頻繁に⽣じていることが明らかになりました。
2. 研究内容と成果
⽪質発⽣期のマウス⼩脳および⼤脳を培養し、DNA ⼆重鎖切断 DSB を標識する蛍光タンパク質※4 を導⼊してニューロン遊⾛の様⼦をライブ観察すると、遊⾛に伴って変形した核内に DSBの形成を⽰すシグナルが現れることが確認されました。発⽣段階を追って解析すると、約 4 割もの遊⾛中のニューロンに DSB の形成が観察されました(図1)。DSB は染⾊体 DNA の複製※5を妨げ、細胞死やガン化を誘導する最も危険な DNA 損傷です。しかし、遊⾛と⽪質形成が完了すると、これらの DSB は消失し、細胞死を起こすことなく脳発⽣が進⾏することがわかりました。
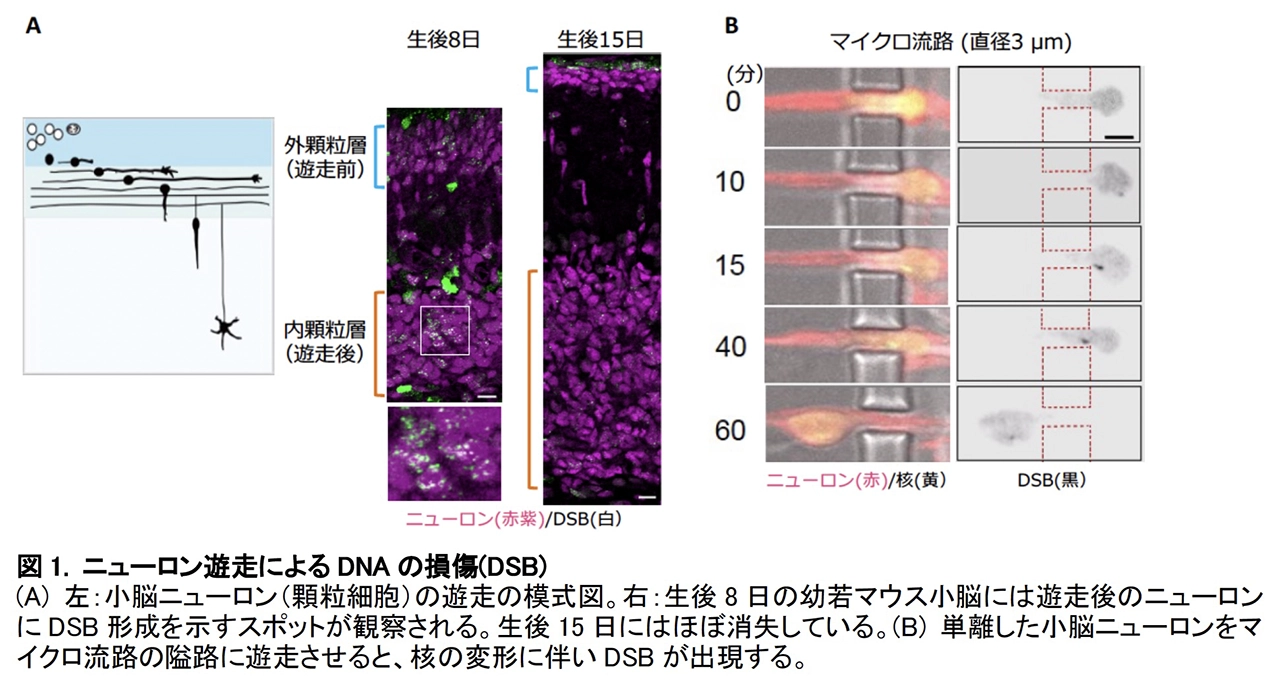
DSB 形成機構を検証するため、脳内の狭⼩空間を再現した、核の直径より⼩さい幅 3 µm(マイクロメートル)のトンネルが連続するマイクロ流路※6 を設計し、単離したニューロンが通過する様⼦をガン細胞株と⽐較して観察しました。その結果、いずれの細胞でも核が隘路通過時に圧迫されると DSB が誘導されることが確認されました(図1B)。また、ガン細胞では核膜が破れ、核質と細胞質が混在する様⼦が観察されました。核膜破損したガン細胞では、核には存在しない細胞質の DNA 分解酵素が核に侵⼊し、⾮特異的に染⾊体 DNA を切断することがわかっています。これに対し、ニューロンでは 3 µm 以下の隘路を遊⾛しても全く核膜の破損が⾒られませんでした。薬理学的解析から、ニューロンの DSB 形成には、DNA にかかる⼒学的ストレスを感知して DNA を切断する核内酵素トポイソメラーゼが関与することがわかりました。さらに次世代シーケンサーを⽤いた解析により、DSB は遺伝⼦本体やその発現制御領域ではなく、機能的影響の少ない不活性なゲノム領域に選択的に蓄積していることが明らかになりました。また、隘路遊⾛で⽣じた DSB はリガーゼ IV というDSB 修復酵素で速やかに修復され、細胞死を起こすことなく正常な状態に戻ることが確認されました(図 2)。
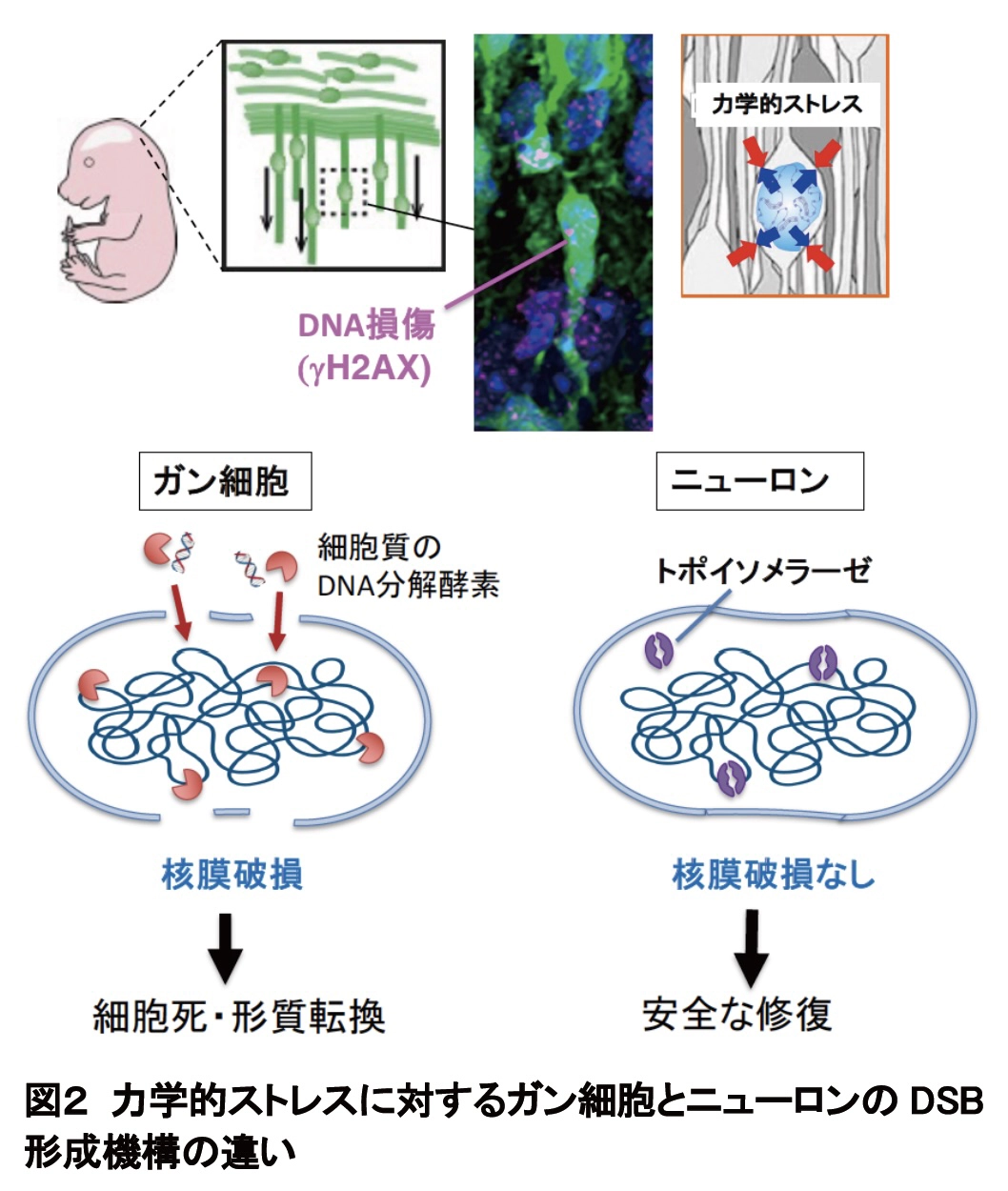
これらの結果から、柔らかい核を持つ遊⾛性細胞が、遊⾛に伴う⼒学的ストレスによる DSB の発⽣が不可避である⼀⽅、ニ ューロンは安全なゲノム領域に DSB を誘発し、速やかに修復して、有害な DSB を残すことなく正常な脳発⽣を完遂するメカニズムを実装していることが⽰唆されます。
さらに、⽪質形成期に多量に⽣じる DSB が修復されなかった場合の影響を検証するため、DSB修復酵素リガーゼ IV をニューロン特異的に⽋損した変異マウスを作成し、個体の脳組織と機能を解析しました(図3A)。変異マウスでは⼩脳ニューロンの遊⾛後も多量の DSB が残存して蓄積しますが、細胞死やシナプス形成※7 に顕著な異常は認められず、個体は外⾒上正常に⽣育しました。しかし、遺伝⼦発現解析の結果、脳内の炎症細胞の活性化およびニューロン機能遺伝⼦の発現低下が認められました(図3B)。そして加齢に伴い、⼩脳機能の低下に起因する運動障害が進⾏しました(図2C)。
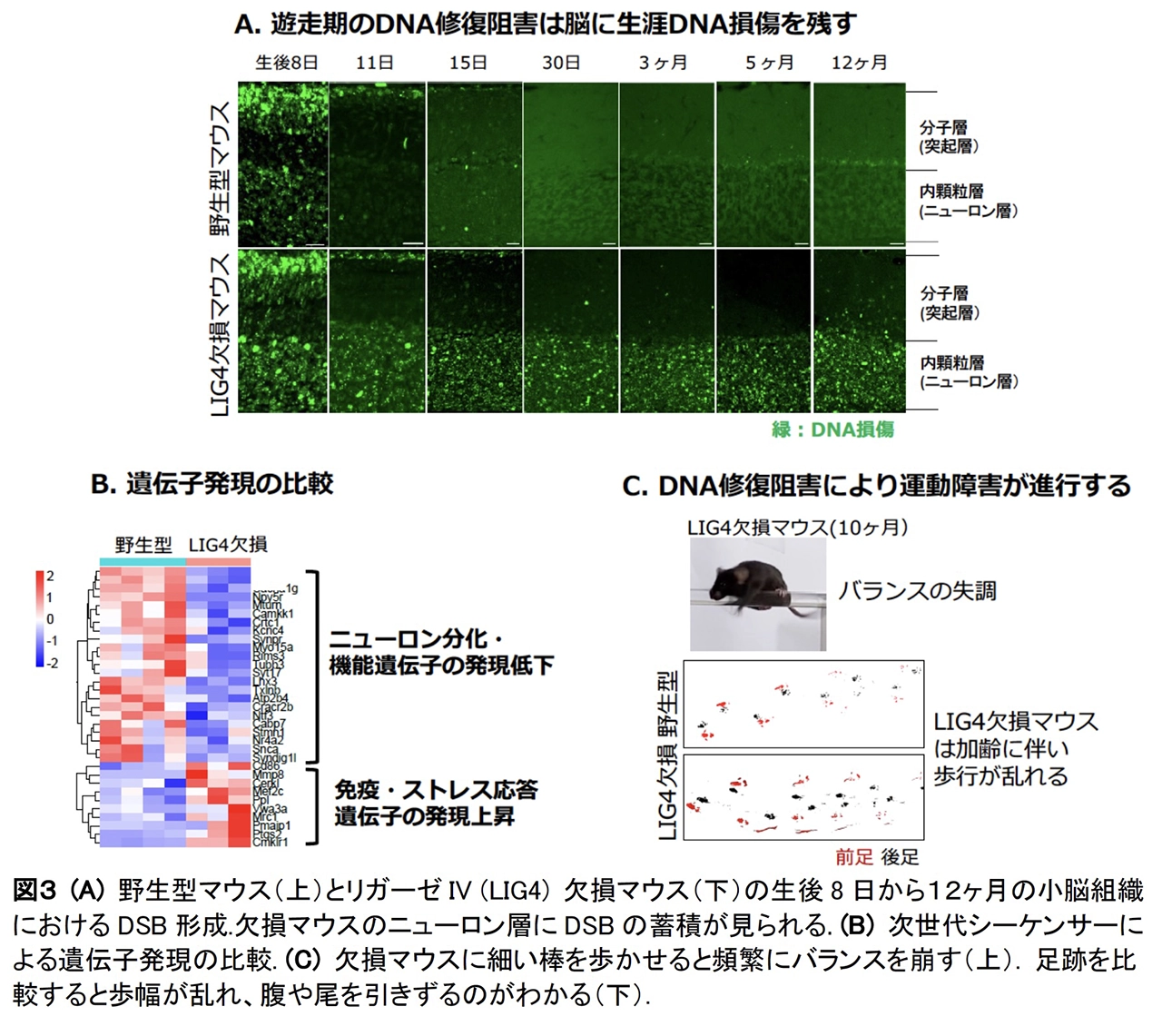
以上の結果から、正常発生過程に⽣じるニューロンの DSBは、通常は適切に制御・修復されるものの、修復エラーが起こると神経機能の低下や疾患発症につながる潜在的リスクであることが明らかになりました。
3. 今後の展開
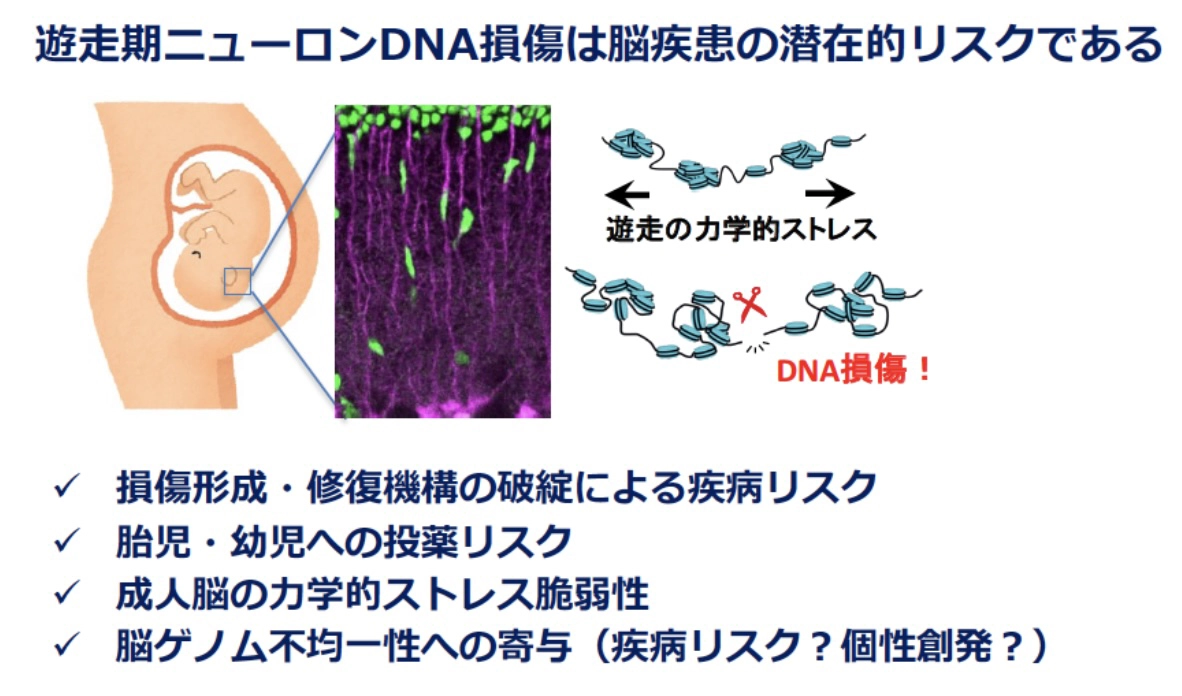
DNA損傷修復の遺伝的な障害は、⽪膚ガンの好発、発育不全、貧⾎などを特徴とするゲノム不安定性疾患を引き起こします。これらの疾患の多くは、進⾏性の⼩脳失調などの神経症状を伴いますが、なぜ⼩脳などの特定の脳領域に症状が現れるのか、その機序は明らかでありませんでした。本研究で発⾒した⽪質形成期の⼀過的DSBは、ゲノム不安定性疾患において修復されずに残存し、神経症状の発症に関与している可能性があります。今後、ゲノム不安定性疾患やその他の発達性障害との関連を精査していく必要があります。
また、DSB 修復に関与するリガーゼ IV は、修復の過程で塩基の⽋失や挿⼊を伴う誤りを⽣じやすく、結果として突然変異を誘発することが知られています。このことから、遊⾛後のニューロンでも⼀定頻度で突然変異が⽣じていると考えられます。ニューロンのゲノムには他の体細胞より多くの突然変異が存在することが知られており、⽪質形成期の DSB がその⼀因である可能性があります。脳に特有のゲノムの特性の理解に向けた新たな⼿掛かりを提供することが期待されます。
用語解説
- ※1 核膜分⼦構造:
- 核膜分⼦構造:核膜はリン脂質の⼆重層でできており、内膜は核ラミナと呼ばれる繊維タンパク質でできた網⽬状構造とそれに結合する凝集したゲノムDNAにより裏打ちされている。核ラミナ分⼦の構成は、核膜の硬さや伸縮性に強く影響する。
- ※2 DNA損傷:
- 環境ストレスや複製などの細胞活動により、DNAの塩基の酸化、脱落、誤った塩基の挿⼊、切断などが起こること。切断には⼀本鎖のみが切れる⼀本鎖切断と⼆重らせんが同時に切れる⼆重鎖切断がある。
- ※3 株化培養細胞:
- 無限に増殖して半永久的に維持することが可能な細胞。ガン組織に由来するものや、遺伝⼦操作で不死化させたものがある。⽣体の細胞の性質や挙動とは必ずしも⼀致しない。
- ※4 蛍光タンパク質:
- 分⼦⽣物学的な⼿法で、特定の波⻑の光を当てると蛍光を発するタンパク質を、標的とするタンパク質と結合させた融合タンパク質を細胞に作らせることにより、蛍光顕微鏡で⽣きた細胞の中の標的タンパク質の挙動を観察することができる。
- ※5 複製:
- 細胞分裂において、⼆本鎖DNAをほどき、それぞれを鋳型として相補鎖を合成して⼆本鎖DNAを倍加するプロセス
- ※6 マイクロ流路:
- ガラスや樹脂を加⼯して微細な溝(ここでは1~5 µm幅)を掘り、液体や細胞が通る流路を形成したデバイス。
- ※7 シナプス形成:
- ニューロン同⼠が神経伝達を⾏うシナプス結合が形成され、神経回路として機能する状態。
研究プロジェクトについて
本研究は、⾰新的先端研究開発⽀援事業AMED-CREST(研究開発課題名:メカノストレスによる脳ゲノム損傷とライフコース疾患リスクの解明)で実施し、⽇本学術振興会・⽂部科学省 科学研究費助成事業(22H05169, 21H00250, 21K19312, 19H04267, JPMXP1323015483, 24K02020, 23K21711, 25K00115, 23K14431)、上原記念⽣命科学財団、武⽥科学振興財団、東京都特別研究費「発がんメカニズム解明と新規がん免疫療法等の研究推進」、⼩野薬品がん・免疫・神経研究財団の助成を受けて⾏われました。