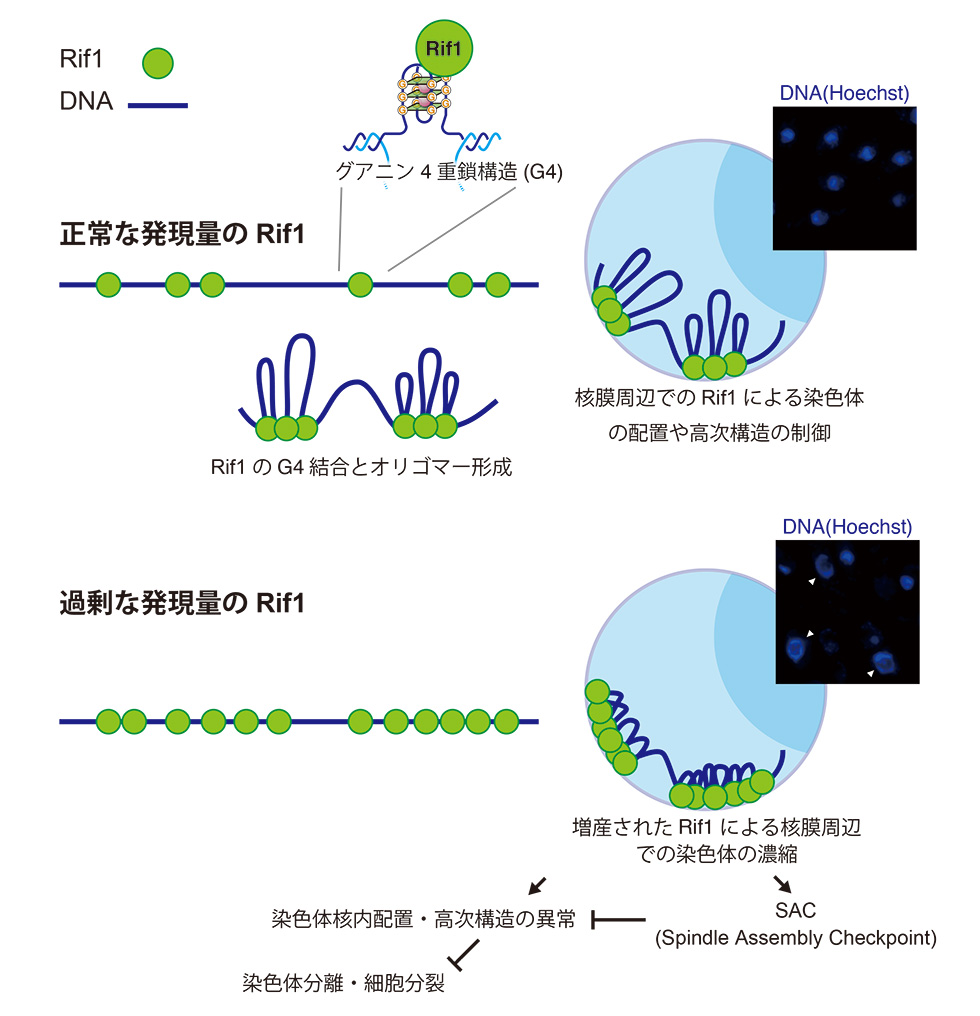
過剰に発現したRif1は染色体を核膜周辺へ異常にリクルートし不均等な染色体分配等を引き起こす
- <論文名>
- “Aberrant association of chromatin with nuclear periphery induced by Rif1 leads to mitotic defect.”
- <発表雑誌>
- Life Science Alliance
DOI: 10.26508/lsa.202201603
- <著者名>
- Kanoh Y., Ueno M., Hayano M., Kudo S., Masai H.
Rif1はDNA複製開始・DNA損傷修復・染色体分離などの制御を介して多くの染色体代謝機能を担っています。我々は、分裂酵母において、Rif1がG-quadruplex (グアニン4重鎖)構造を認識し染色体へ結合し、その多量体形成とDNA結合を介して染色体高次構造を形成し、複製開始のタイミングを制御することを見出しました。
1. Rif1の増産は、S期以外のイベントの阻害を介して細胞死を誘導する
動物細胞Rif1は、細胞種によりその発現量が異なっており、これが、細胞特異的な核内の染色体構造に影響を与えると考えられます。今回、私たちはRif1の発現量の変化が核内染色体構造や細胞増殖・細胞周期にどのように影響するか分裂酵母で検証しました。Rif1の過剰な発現は細胞形態の異常と細胞死を引き起こしました。細胞周期の解析から、Rif1増産はDNA複製を阻害することが示されました。しかし、PP1(脱リン酸化酵素)結合部位変異体は複製を阻害しませんでしたが、細胞死を誘導することから、DNA複製抑制ではなく、それ以外の細胞周期のイベントの阻害が細胞死の原因であることが示唆されました。
2. Rif1の増産は、分裂期の異常、染色体の不均等分配を誘導する。
細胞の形態の観察から、染色体の不均等分配等が起こっており、染色体の分配を担うチューブリンの伸長が停止している細胞が多く観察されました。この現象は、染色体分配チェックポントであるSpindle Assembly Checkpoint (SAC)が活性化されている可能性を示唆していました。そこで、SACに必要とされるMad2もしくはBub1の欠損株にRif1の過剰発現を誘導すると、より強い細胞死が引き起こされました。これは、Rif1過剰発現による染色体分配異常をSACが一部抑制していることを示します。
3. Rif1の増産は、染色体を核膜に集積させる。
染色体の核内の配置を注意深く観察すると、Rif1の過剰発現を誘導した細胞で染色体が核膜周辺に濃縮されていることに気づきました。私たちは哺乳類細胞でRif1は染色体を核膜周辺に繋ぎ止める機能があることを見出していました。Rif1の過剰発現が誘導され染色体を異常に核膜周辺に繋ぎ止められ、分配時に染色体の正常に行われなかった可能性が示されました。また、Rif1の染色体への結合部位を決定する手法であるChIP-seq解析をRif1過剰発現細胞で行ったところ、Rif1の染色体への結合部位が増加していました。一方、染色体結合能が欠失したRif1の過剰発現は、染色体の異常な核膜周辺への移動や細胞死を誘導しませんでした。
4. 結論
これらの結果から、Rif1が過剰発現すると、その染色体結合能に依存して、染色体を核膜近傍に蓄積させ、SACを誘導するとともに、染色体の均等な分配を阻害し、最終的に細胞死を誘導すると考えられます。これらの事実は、適切なRif1の発現が染色体の核内での配置調整に重要であることを示しています。
5. 発見の意義
染色体の核内配置や、高次構造は、遺伝子発現や複製に大きな影響を及ぼします。その異常は、ゲノムの不安定性を誘導し、がんやその他の疾患発生の原因となります。Rif1は染色体の核内配置や、高次構造の制御によりゲノム複製や修復を制御する重要な因子です。今回の発見は、Rif1の発現異常により染色体の核内配置や、高次構造に異常をきたし、ゲノム不安定性の最も重要な要因である染色体分配過程に障害をもたらすことを示し、疾患発生の原因となる染色体異常の誘導のメカニズムに重要な知見をもたらすものです。