
フォスファカンはTAG-1/GD3ラフトに依存する小脳顆粒細胞の反発因子として働く
細胞膜研究室の小松谷啓介研究員、笠原浩二研究室長らと、長岡技術科学大学の霜田靖准教授、鹿児島大学の武田泰生教授、東京理科大学の下仲基之教授、愛知医科大学の杉浦信夫客員研究員、神経回路形成プロジェクト前田信明リーダーらとの共同研究グループは、コンドロイチン硫酸プロテオグリカンであるフォスファカンが小脳顆粒細胞の反発因子として働く仕組みについて解明しました。
本研究は、国際学術誌「Journal of Neurochemistry」(2022年10月13日)に掲載されました。
- <論文名>
- “Phosphacan acts as a repulsive cue in murine and rat cerebellar granule cells in a TAG-1/GD3 rafts-dependent manner”
- <著者名>
- Keisuke Komatsuya, Tomohiro Iguchi, Michiko Fukuyama, Ikuo Kawashima, Kiyoshi Ogura, Norihito Kikuchi, Yasushi Shimoda, Yasuo Takeda, Motoyuki Shimonaka, Naomasa Yamamoto, Nobuo Sugiura, Nobuaki Maeda, Kohji Kasahara
- <発表雑誌>
- Journal of Neurochemistry
DOI:10.1111/jnc.15709
背景
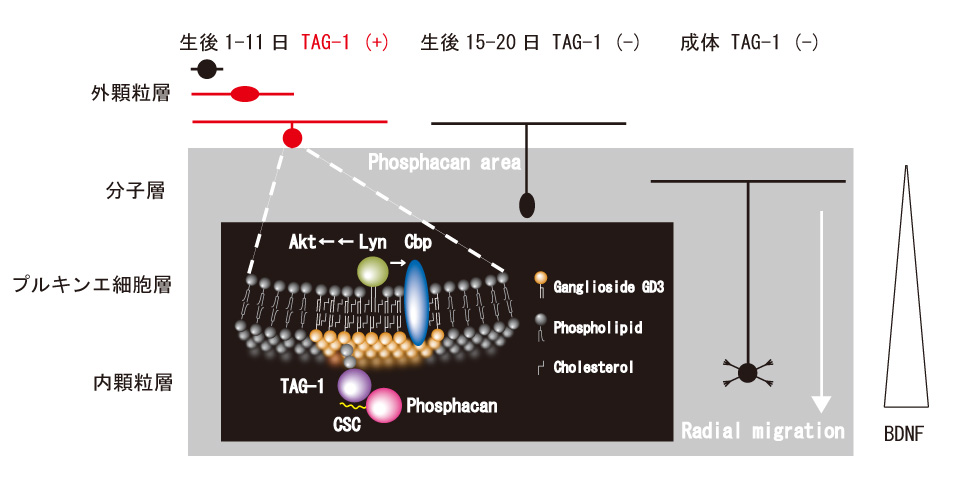
コンドロイチン硫酸プロテオグリカンであるフォスファカンは、小脳顆粒細胞の反発因子として働くことをこれまでに報告しましたが、その反発機構は不明でした。フォスファカンは、軸索伸長や細胞移動活性を持つGPIアンカー型神経細胞接着分子TAG-1とin vitroの系でコンドロイチン硫酸依存的に相互作用することが知られていたことから、TAG-1欠損マウスの小脳顆粒細胞に対するフォスファカンの反発を解析しました。すると反発作用がほとんどなかったことから、TAG-1はフォスファカンの受容体として働いていると考えられました。表面プラズモン共鳴解析によりTAG-1と相互作用するのはフォスファカンのコンドロイチン硫酸の一つであるCSC(コンドロイチン6硫酸)であることが分かりました。顆粒細胞は生後1日目から20日目にかけて外顆粒層で増殖、分子層、プルキンエ細胞層を通りぬけ内顆粒層に移動します。生後1日から11日までTAG-1は一過性に外顆粒層の顆粒細胞にのみ局在し、フォスファカンは隣接する分子層から内顆粒層にかけて存在し、TAG-1染色と接していました。顆粒細胞の外顆粒層から内顆粒層への移動は、内顆粒層に局在するBDNF(Brain-derived neurotrophic factor)による誘因作用によると考えられています。そこでBoyden chamberを使った細胞移動実験系で、BDNFによる顆粒細胞の移動がフォスファカンおよびCSCで抑制されることを見出しました。我々はこれまでに、フォスファカンが小脳顆粒細胞のGD3ラフトにおいてTAG-1を介するシグナルを伝達していることを報告してきました。これらのことから、フォスファカンのTAG-1/GD3ラフトを介する反発作用により顆粒細胞の外顆粒層からの移動が妨げられ、TAG-1の消失に伴い移動が可能になると考えられました。
<用語解説>
- コンドロイチン硫酸プロテオグリカン
- グルクロン酸とNアセチルガラクトサミンの二糖ユニットが繰返した糖鎖であるコンドロイチン硫酸とコアタンパク質が共有結合したハイブリッド分子。分子間相互作用を介して細胞外マトリックスを構成して、神経系においては軸索ガイダンスの調節因子として働く。
- GPIアンカー
- グリコシルホスファチジルイノシトール(GPI)アンカーの略で、細胞膜にタンパク質を結合させる糖脂質性のアンカー(錨=いかり)のこと。GPIを結合し、これをアンカーとして膜につなぎ止められている一群のタンパク質をGPIアンカー型タンパク質という。
- ラフト
- スフィンゴ糖脂質、スフィンゴミエリン、コレステロールに富む細胞膜上に存在するミクロドメインである。GPIアンカー型タンパク質や様々なシグナル伝達分子を結合させることにより、細胞膜を介するシグナル伝達の中継点として働き、多くの生命現象を調節している。